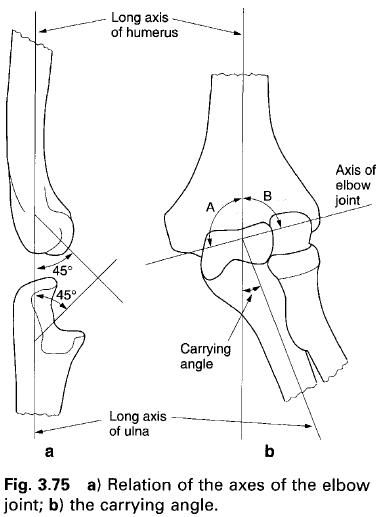
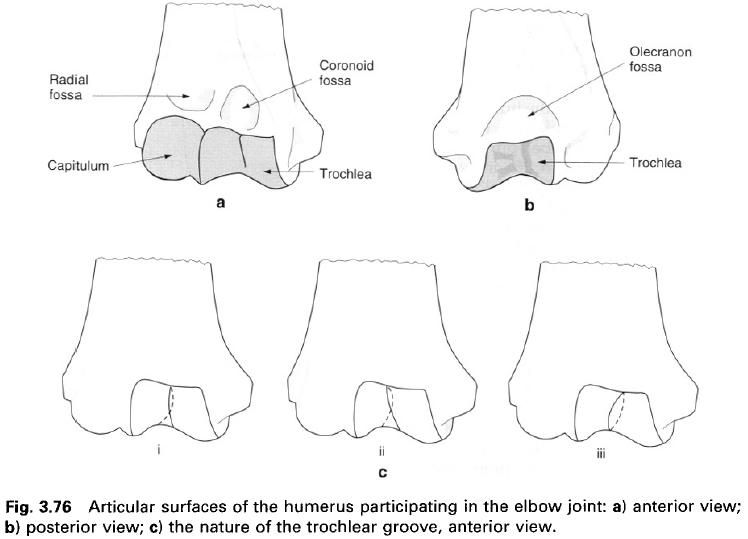
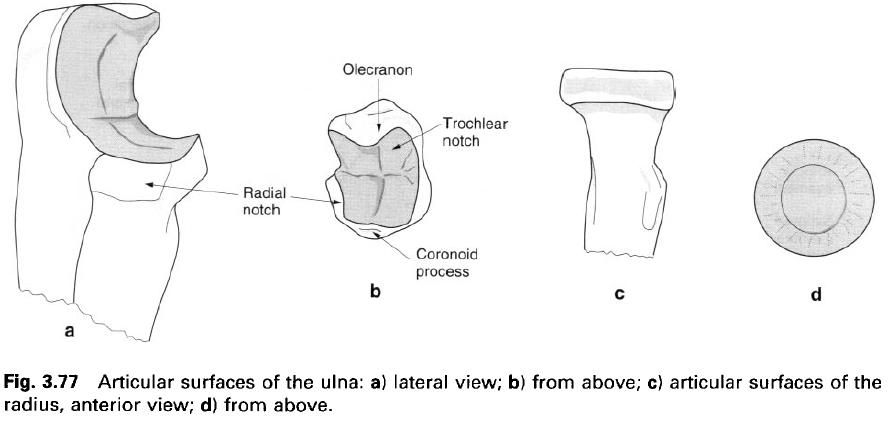
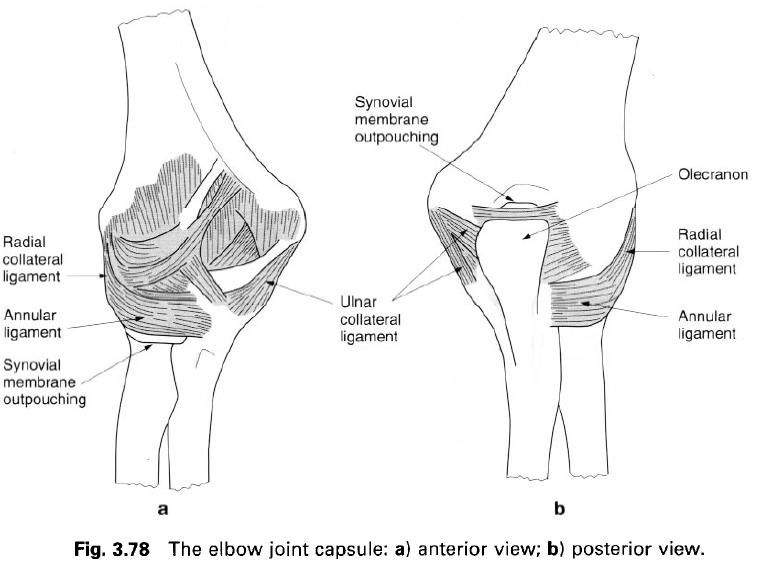
I figured we should start this myth destroying article off with the most
annoying myth first. Lord, when will this one go away? Now the average
reader person is probably thinking "who in the world still believes
that ridiculous statement?" The answer is a great deal of people, even
well educated medical professionals and scientists who should know better,
still believe this to be true. Don't forget, the
high carb,
low fat,
low protein diet recommendations are alive and well with the average
nutritionist, doctor, and of course the "don't confuse us with the
facts" media following close behind.
For the past half century or so scientists using crude methods and poor
study design with sedentary people have held firm to the belief that
bodybuilders, strength athletes of various types, runners, and other highly
active people did not require any more protein than Mr. Potato
Head.....err, I mean the average couch potato. However, In the past few
decades researchers using better study designs and methods with real live
athletes have come to a different conclusion altogether, a conclusion hard
training bodybuilders have known for years. The fact that active people do
indeed require far more protein than the RDA to keep from losing hard
earned muscle tissue when dieting or increasing muscle tissue during the
off season.
In a recent review paper on the subject one of the top researchers in the
field (Dr. Peter Lemon) states "...These data suggest that the RDA for
those engaged in regular endurance exercise should be about 1.2-1.4 grams
of protein/kilogram of body mass (150%-175% of the current RDA) and 1.7 -
1.8 grams of protein/kilogram of body mass per day (212%-225% of the
current RDA) for strength exercisers."
Another group of researchers in the field of
protein metabolism have come
to similar conclusions repeatedly. They found that strength training
athletes eating approximately the RDA/RNI for protein showed a decreased
whole body protein synthesis (losing muscle jack!) on a protein intake of
0.86 grams per kilogram of bodyweight. They came to an almost identical
conclusion as that of Dr. Lemon in recommending at least 1.76g per kilogram
of bodyweight per day for strength training athletes for staying in
positive nitrogen balance/increases in whole body
protein synthesis.
This same group found in later research that endurance athletes also need
far more
protein than the RDA/RNI and that men catabolize (break down) more
protein than women during endurance exercise.
They concluded "In summary,
protein requirements for athletes
performing strength training are greater than sedentary individuals and are
above the current Canadian and US recommended daily protein intake requirements
for young healthy males." All I can say to that is, no sh%# Sherlock?!
Now my intention of presenting the above quotes from the current research
is not necessarily to convince the average athlete that they need more
protein than Joe shmoe couch potato, but rather to bring to the readers
attention some of the figures presented by this current research. How does
this information relate to the eating habits of the average athlete and the
advice that has been found in the lay bodybuilding literature years before
this research ever existed? With some variation, the most common advice on
protein intakes that could be-and can be- found in the bodybuilding
magazines by the various writers, coaches, bodybuilders, etc., is one gram
of protein per pound of body weight per day.
So for a 200 pound guy that would be 200 grams of protein per day. No
sweat. So how does this advice fair with the above current research
findings? Well let's see. Being scientists like to work in kilograms (don't
ask me why) we have to do some converting. A kilogram weighs 2.2lbs. So,
200 divided by 2.2 gives us 90.9. Multiply that times 1.8 (the high end of
Dr. Lemon's research) and you get 163.6 grams of protein per day. What
about the nutritionists, doctors, and others who call(ed) us "protein
pushers" all the while recommending the RDA as being adequate for
athletes?
Lets see. The current RDA is 0.8 grams of protein per kilogram of
bodyweight: 200 divided by 2.2 x 0.8 = 73 grams of
protein per day for a
200lb person. So who was closer, the bodybuilders or the arm chair
scientists? Well lets see! 200g (what bodybuilders have recommended for a
200lb athlete) - 163g ( the high end of the current research
recommendations for a 200lb person) = 37 grams (the difference between what
bodybuilders think they should eat and the current research).
How do the RDA pushers fair? Hey, if they get to call us "protein
pushers" than we get to call them "RDA pushers!" Anyway,
163g - 73g = (drum role) 90 grams! So it would appear that the bodybuilding
community has been a great deal more accurate about the
protein needs of
strength athletes than the average nutritionist and I don't think this
comes as any surprise to any of us. So should the average bodybuilder
reduce his protein intake a bit from this data? No, and I will explain why.
As with vitamins and other nutrients, you identify what looks to be the
precise amount of the compound needed for the effect you want (in this case
positive nitrogen balance, increased protein synthesis, etc) and add a
margin of safety to account for the biochemical individuality of different
people, the fact that there are low grade protein sources the person might
be eating, and other variables.
So the current recommendation by the majority of bodybuilders, writers,
coaches, and others of one gram per pound of bodyweight does a good job of
taking into account the current research and adding a margin of safety. One
things for sure, a little too much protein is far less detrimental to the
athletes goal(s) of increasing muscle mass than too little protein, and
this makes the RDA pushers advice just that much more.... moronic, for lack
of a better word.
There are a few other points I think are important to look at when we
recommend additional protein in the diet of athletes, especially strength
training athletes. In the off season, the strength training athletes needs
not only adequate protein but adequate calories. Assuming our friend (the
200lb bodybuilder) wants to eat approximately 3500 calories a day, how is
he supposed to split his calories up? Again, this is where the bodybuilding
community and the conservative nutritional/medical community are going to
have a parting of the ways... again. The conservative types would say
"that's an easy one, just tell the bodybuilder he should make up the
majority of his calories from carbohydrates."
Now lets assume the bodybuilder does not want to eat so many
carbs. Now the
high carb issue is an entirely different fight and article, so I am just
not going to go into great depth on the topic here. Suffice it to say,
anyone who regularly reads articles, books, etc, >from people such as
Dan Duchaine, Dr. Mauro Dipasquale, Barry Sears PhD, Udo Erasmus PhD, yours
truly, and others know why the high carb diet bites the big one for losing
fat and gaining muscle (In fact, there is recent research that suggests
that carbohydrate restriction, not calorie restriction per se, is what's
responsible for mobilizing fat stores). So for arguments sake and lack of
space, let's just assume our 200lb bodybuilder friend does not want to eat
a high carb diet for his own reasons, whatever they may be.
What else can he eat? He is only left with fat and protein. If he splits up
his diet into say 30% protein, 30 % fat, and 40% carbs, he will be eating 1050
calories as protein (3500x30% = 1050) and 262.5g of protein a day (1050
divided by 4 = 262.5). So what we have is an amount (262.5g) that meets the
current research, has an added margin of safety, and an added component for
energy/calorie needs of people who don't want to follow a high carb diet,
hich is a large percentage of the bodybuilding/strength training community.
here are other reasons for a high protein intake such as hormonal effects
(i.e. effects on IGF-1, GH, thyroid ), thermic effects, etc., but I think I
have made the appropriate point. So is there a time when the bodybuilder
might want to go even higher in his percent of calories >from protein
than 30%? Sure, when he is dieting.
It is well established that carbs are "protein sparing" and so
more protein is required as percent of calories when one reduces calories.
Also, dieting is a time that preserving lean mass (muscle) is at a premium.
Finally, as calories decrease the quality and quantity of protein in the
diet is the most important variable for maintaining muscle tissue (as it
applies to nutritional factors), and of course protein is the least likely
nutrient to be converted to bodyfat. In my view, the above information
bodes well for the high protein diet. If you tell the average RDA pusher
you are eating 40% protein while on a diet, they will tell you that 40% is
far too much protein. But is it? Say our 200lb friend has reduced his
calories to 2000 in attempt to reduce his bodyfat for a competition, summer
time at the beach, or what ever. Lets do the math. 40% x 2000 = 800
calories from protein or 200g (800 divided by 4). So as you can see, he is
actually eating less protein per day than in the off season but is still in
the range of the current research with the margin of safety/current
bodybuilding recommendations intact.
Bottom line? High protein diets are far better for reducing bodyfat,
increasing muscle mass, and helping the hard training bodybuilder achieve
his (or her!) goals, and it is obvious that endurance athletes will also
benefit from diets higher in protein than the worthless and outdated RDAs.
Myth #2 "High protein diets are bad for you"
So the average person reads the above information on the protein needs and
benefits of a high protein diet but remembers in the back of their mind
another myth about high protein intakes. "I thought high protein diets
are bad for the kidneys and will give you osteoporosis! " they exclaim
with conviction and indignation. So what are the medical facts behind these
claims and why do so many people, including some medical professionals and
nutritionists, still believe it?
For starters, the negative health claims of the high protein diet on kidney
function is based on information gathered from people who have preexisting
kidney problems. You see one of the jobs of the kidneys is the excretion of
urea (generally a non toxic compound) that is formed from ammonia (a very
toxic compound) which comes from the protein in our diets. People with
serious kidney problems have trouble excreting the urea placing more stress
on the kidneys and so the logic goes that a high protein diet must be hard
on the kidneys for healthy athletes also.
Now for the medical and scientific facts. There is not a single scientific
study published in a reputable peer - reviewed journal using healthy adults
with normal kidney function that has shown any kidney dysfunction what so
ever from a high protein diet. Not one of the studies done with healthy
athletes that I mentioned above, or other research I have read, has shown
any kidney abnormalities at all. Furthermore, animals studies done using
high protein diets also fail to show any kidney dysfunction in healthy
animals.
Now don't forget, in the real world, where millions of athletes have been
following high protein diets for decades, there has never been a case of
kidney failure in a healthy athlete that was determined to have been caused
solely by a high protein diet. If the high protein diet was indeed putting
undo stress on our kidneys, we would have seen many cases of kidney
abnormalities, but we don't nor will we. From a personal perspective as a
trainer for many top athletes from various sports, I have known
bodybuilders eating considerably more than the above research recommends
(above 600 grams a day) who showed no kidney dysfunction or kidney problems
and I personally read the damn blood tests! Bottom line? 1-1.5 grams or
protein per pound of bodyweight will have absolutely no ill effects on the
kidney function of a healthy athlete, period. Now of course too much of
anything can be harmful and I suppose it's possible a healthy person could
eat enough protein over a long enough period of time to effect kidney
function, but it is very unlikely and has yet to be shown in the scientific
literature in healthy athletes.
So what about the osteoporosis claim? That's a bit more complicated but the
conclusion is the same. The pathology of osteoporosis involves a
combination of many risk factors and physiological variables such as macro
nutrient intakes (carbs, proteins, fats), micro nutrient intakes (vitamins,
minerals, etc), hormonal profiles, lack of exercise, gender, family
history, and a few others. The theory is that high protein intakes raise
the acidity of the blood and the body must use minerals from bone stores to
"buffer" the blood and bring the blood acidity down, thus
depleting one's bones of minerals. Even if there was a clear link between a
high protein diet and osteoporosis in all populations (and there is not)
athletes have few of the above risk factors as they tend to get plenty of
exercise, calories, minerals, vitamins, and have positive hormonal
profiles. Fact of the matter is, studies have shown athletes to have denser
bones than sedentary people, there are millions of athletes who follow high
protein diets without any signs of premature bone loss, and we don't have
ex athletes who are now older with higher rates of osteoporosis.
In fact, one recent study showed women receiving extra protein from a
protein supplement had increased bone density over a group not getting the
extra protein! The researchers theorized this was due to an increase in
IGF-1 levels which are known to be involved in bone growth. Would I
recommend a super high protein diet to some sedentary post menopausal
woman? Probably not, but we are not talking about her, we are talking about
athletes. Bottom line? A high protein diet does not lead to osteoporosis in
healthy athletes with very few risk factors for this affliction, especially
in the ranges of protein intake that have been discussed throughout this
article.
Myth #3 "All proteins are created equal"
How many times have you heard or read this ridiculous statement? Yes, in a
sedentary couch potato who does not care that his butt is the same shape as
the cushion he is sitting on, protein quality is of little concern.
However, research has shown repeatedly that different proteins have various
functional properties that athletes can take advantage of. For example,
whey protein concentrate (WPC) has been shown to improve immunity to a variety
of challenges and intense exercise has been shown to compromise certain
parts of the immune response. WPC is also exceptionally high in the branch
chain amino acids which are the amino acids that are oxidized during
exercise and have been found to have many benefits to athletes. We also
know soy has many uses for athletes.
Anyway, I could go on all day about the various functional properties of
different proteins but there is no need. The fact is that science is
rapidly discovering that proteins with different amino acid ratios (and
various constituents found within the various protein foods) have very
different effects on the human body and it is these functional properties
that bodybuilders and other athletes can use to their advantage. Bottom
line? Let the people who believe that all proteins are created equal
continue to eat their low grade proteins and get nowhere while you laugh
all the way to a muscular, healthy, low fat body!
Conclusion
Over the years the above myths have been floating around for so long they
have just been accepted as true, even though there is little to no research
to prove it and a whole bunch of research that disproves it! I hope this
article has been helpful in clearing up some of the confusion for people
over the myths surrounding protein and athletes. Of course now I still have
to address even tougher myths such as "all fats make you fat and are
bad for you," "supplements are a waste of time," and my personal
favorite, "a calorie is a calorie." The next time someone gives
you a hard time about your high protein intake, copy the latest study on
the topic and give it to em. If that does not work, role up the largest
bodybuilding magazine you can find and hit them over the head with it!