Aerobic
training, or cardiorespiratory endurance training, improves central and peripheral blood flow
and enhances the capacity of the muscle fibers to generate greater amounts of adenosine triphosphate(ATP).
Endurance: Muscular versus cardiorespiratory
Endurance is a term that refers to two separate
but related concepts: muscular endurance and cardiorespiratory endurance. Each
makes a unique contribution to athletic performance, and each differs in its
importance to different athletes.
For sprinters, endurance is the quality that
allows them to sustain a high speed over the full distance of, for example, a
100 or 200m race. This component of fitness is termed muscular endurance, the
ability of a single muscle or muscle group to sustain high-intensity,
repetitive, or static exercise. This type of endurance is also exemplified by
the weightlifter doing multiple repetitions, the boxer, and the wrestler. The
exercise or activity can be rhythmic and repetitive in nature, such as multiple
repetitions of the bench press for the weightlifter and jabbing for the boxer.
Or the activity can be more static, such as a sustained muscle action when a
wrestler attempts to pin an opponent to the mat. In either case, the resulting
fatique is confined to a specific muscle group, and the activity duration is
usually no more than 1 to 2 min. Muscular endurance is highly related to
muscular strength and to anaerobic power development.
While muscular endurance is specific to
individual muscles or muscle groups, cardiorespiratory endurance relates to the
entire body’s ability to sustain prolonged, dynamic exercise using large muscle
groups. This type of endurance is used by the cyclist, distance runner, or
endurance swimmer who completes long distances at a fairly fast pace.
Cardiorespiratory endurance is related to the development of the cardiovascular
and respiratory systems’ ability to maintain oxygen delivery to working muscles
during prolonged bouts of exercise, as well as the muscles’ ability to utilize
energy aerobically. This is why the terms cardiorespiratory endurance and
aerobic endurance are sometimes used synonymously.
Evaluating
cardiorespiratory endurance capacity
To study training effects on endurance, there
needs to be an objective, repeatable means of measuring an individual’s
cardiorespiratory endurance capacity. In that way, the exercise scientist,
coach, or athlete can monitor improvements as physiological adaptations occur
during the training program.
Maximal endurance capacity: VO2max or aerobic power
Most exercise scientists regard VO2max, sometimes called
maximal aerobic power or maximal aerobic capacity, as the best objective
laboratory measure of maximal cardiorespiratory endurance. VO2max is
defined as the highest rate of oxygen consumption attainable during maximal or
exhaustive exercise. VO2max is defined as the highest rate of oxygen
consumption attainable during maximal or exhaustive exercise. VO2max
as defined by the Fick equation is
dictated by maximal cardiac output(delivery
of oxygen and blood flow to working muscles) and the maximal(a-ṽ)O2
difference(the ability of the active muscles to extract and use the oxygen). As
exercise intensity increases, oxygen consumption eventually either plateaus or
decreases slightly, even with further increases in workload, indicating that a
truly maximal VO2 has been achieved.
With endurance training, more oxygen can be
delivered to, and consumed by, active muscles than in an untrained state.
Previously untrained subjects demonstrate average increases in VO2max
of 15% to 20% after a 20-week training program. These improvements allow
individuals to perform endurance activities at a higher intensity, improving
their performance potential. Figure below illustrates the increase of VO2max
after 12 months of aerobic training in a previously untrained individual. In
this example, VO2max increased by about 30%. Note that the VO2
“cost” of running at a certain submaximal intensity did not change but that
higher running speeds could be attained after training.

Submaximal
endurance capacity
In addition to increasing maximal endurance
capacity, endurance training increases submaximal endurance capacity, which is
much more difficult to evaluate. Steady-state submaximal heart rate at the same
exercise intensity measured before and after training is one physiological
variable that can be used to objectively quantify the effect of training. Additionally,
exercise scientists have used performance measures to quantify submaximal
endurance capacity. For example, one test used to determine submaximal
endurance capacity is the average peak absolute power output a person can
maintain over a fixed period of time on a cycle ergometer. For running, the
average peak speed of velocity a person can maintain during a fixed period of
time would be similar type of test. Generally, these tests will last 30 min to
an hour.
Submaximal endurance capacity is more closely
related to actual competitive endurance performance than VO2max, and
is likely determined by both the person’s VO2max and the threshold
for his or her onset of blood lactic acid accumulation(OBLA) – that point at which lactate begins to appear
at a disproportionate rate in the blood. With endurance training, submaximal
endurance capacity increases.
Cardiovascular adaptations to training
Numerous cardiovascular adaptations occur in
response to exercise training, including changes in the following
cardiovascular variables:
- Heart size
- Stroke volume
- Heart rate
- Cardiac output
- Blood flow
- Blood pressure
- Blood volume.
To understand completely, it is very important
to review how these components relate to oxygen transport.
Oxygen
transport system
Cardiorespiratory endurance is related to the
cardiovascular and respiratory systems’
ability to deliver sufficient oxygen to meet the needs of metabolically
active tissues.
The ability of the cardiovascular and
respiratory systems to deliver oxygen to active tissues is defined by Fick equation. The Fick equation states that systemic oxygen consumption is
determined by both the delivery of oxygen(cardiac output) via blood flow and
the amount of oxygen extracted by the tissues, the (a-ṽ)O2
difference. The product of cardiac output and the (a-ṽ)O2
difference determines the rate of which oxygen is being consumed:
VO2 = stroke volume x heart rate x
(a-ṽ)O2 diff.
The oxygen demand of exercising muscles
increases with increasing exercise intensity. Aerobic endurance depends on the
cardiorespiratory system’s ability to deliver sufficient oxygen to these active
tissues to meet their heightened demands for oxygen for oxidative metabolism.
As maximal levels of exercise are achieved, heart size, blood flow, blood
pressure, and blood volume can all potentially limit the maximal ability to
transport oxygen. Endurance training elicits numerous changes in these
components of the oxygen transport system that enable it to function more
effectively.
Heart size
As an adaptation to the increased work demand, heart mass and volume increase with
training. Cardiac muscle, like skeletal muscle, undergoes morphological
adaptations as a result of chronic endurance training. At one time, cardiac hypertrophy induced by exercise
– “athlete’s heart”, as it was called – caused
some concern because experts incorrectly believed that enlargement of the heart
always reflected a pathological state, as sometimes occurs with severe
hypertension. Training-induced cardiac hypertrophy is now recognized as a
normal adaptation to chronic endurance training.
The left ventricle, does the most work and thus
undergoes the greatest adaptation in response to endurance training. The extent
and location of heart size adaptations depend on the type of exercise training
performed. For example, during resistance training, the left ventricle must
contract against increased afterload from the systemic circulation. It was
postulated to overcome this high afterload, the heart muscle compensated by
increasing left ventricular wall thickness, thereby increasing its
contractility. Blood pressure during resistance training can exceed 480/350
mmHg. This presents a considerable resistance that must be overcome by the left
ventricle. Thus, the increase in its muscle mass is in direct response to
repeated exposure to the increased afterload with resistance training.
With endurance training, left ventricular
chamber size increases. This allows for increased left ventricular filling and
consequently an increase in stroke volume. The increase in left ventricular
dimensions is largely attributable to a training-induced increase in plasma
volume that increases left ventricular end-diastolic volume(increased preload).
In concert with this, a decrease in heart rate at rest caused by increased
parasympathetic tone, and during exercise at the same rate of work, allows a
longer diastolic filling period. The increases in plasma volume and diastolic
filling time increase left ventricular chamber size at the end of diastole.
It was originally hypothesized that this
increase in left ventricular dimensions was the only change in the left
ventricle caused by endurance training. Additional research has revealed that
myocardial wall thickness also increases with endurance training, rather than
just with resistance training. Using magnetic resonance imaging, Milliken and
colleagues found that highly trained endurance athletes(competitive
cross-country skiers, endurance cyclists, and long-distance runners) had
greater left ventricular masses than did non-endurance-trained control
subjects. Left ventricular mass was highly correlated with VO2max or
aerobic power.
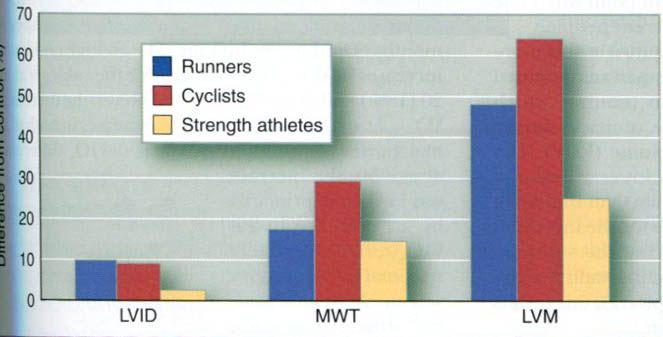
Fagard conducted the most extensive review of
the existing research literature in 1996, focusing on long-distance runners(135
athletes and 173 controls), cyclists(69 athletes and 65 controls), and strength
athletes(178 athletes, including weight- and powerlifters, bodybuilders,
wrestlers, throwers, and bobsledders, and 105 controls). For each group, the
athletes were matched by age and body size with a group of sedentary control
subjects. For each group of runners, cyclists, and strength athletes, the
internal diameter of the left ventricle(LVID, an index of chamber size) and the
total left ventricular mass(LVM) were greater in the athletes compared with
their age- and sized-matched controls(figure below). Thus, data from this large
cross-sectional study support the hypothesis that both left ventricular chamber
size and wall thickness increase with endurance training.
Most studies of the heart size changes with
training have been cross-sectional, comparing trained individuals with
sedentary, untrained individuals. We can learn much from cross-sectional
studies, but they do not provide us with the same information that we could get
from studying a group of untrained people who train for months or years,
focusing on their changes from the untrained to the trained state. Certainly a
portion of the differences that we see in figure above us can be attributed to
genetics, not training. However, a number of studies have followed individuals
from an untrained state to a trained state to an untrained state. These studies
have reported increases in heart size with training and decreases with
detraining. So, training does bring about changes, but they are likely not as
large as the differences we see in the figure above us.
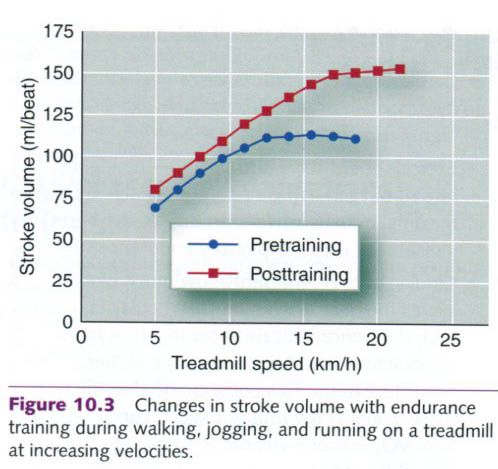
Stroke volume
As a result of endurance training, stroke
volume increases. Stroke volume
at rest is substantially higher after an endurance training program than it
is before training. This endurance training-induced increase is also seen at a
given submaximal exercise intensity and at maximal exercise. This increase
is illustrated in figure below, which shows the changes in stroke volume of a
subject who exercised at increasing intensities up to maximal intensity before
and after a six-month aerobic endurance training program. Typical values for
stroke volume at rest and during maximal exercise in untrained, trained, and
highly trained athletes are listed in the table below. The wide variation in
stroke volume values for any given cell within this table is largely
attributable to differences in body size. Absolute stroke volume at rest and
during exercise is not merely a function of a person’s state of training but
also reflects differences in body size. Larger people typically have larger
hearts and a greater blood volume, and thus higher stroke volumes – an
important point when one is comparing stroke volumes of different people.
|
Stroke
volumes(SV) for different states of training
|
||
|
Subjects
|
SVrest(ml/beat)
|
SVmax(ml/beat)
|
|
Untrained
|
50-70
|
80-110
|
|
Trained
|
70-90
|
110-150
|
|
Highly
trained
|
90-110
|
150- >220
|
After aerobic training, the left ventricle
fills more completely during diastole than it does in an untrained state.
Plasma volume expands with training, which allows for more blood to enter the
ventricle during diastole, increasing end-diastolic volume(EDV). The heart rate of a trained heart is also
lower at rest and at the same absolute exercise intensity than that of an
untrained heart, allowing more time for the increased diastolic filling. More
blood entering the ventricle increases the stretch on the ventricular walls;
by the Frank-Starling mechanism, this results in an increase in force of
contraction.
The thickness of the posterior and septal walls
of the left ventricle also increases slightly with endurance training.
Increased ventricular muscle mass results in increased contractile force, in
turn causing end-systolic volume to decrease. More blood is forced out of the
heart, leaving less blood in the left ventricle after systole.
The decrease in end-systolic volume is
augmented by the decrease in peripheral resistance that occurs with training.
Increased contractility resulting from an increase in left ventricular
thickness and greater diastolic filling(Frank-Starling mechanism), coupled with the reduction in systemic peripheral resistance,
increases the ejection fraction[equal to (EDV-ESV)/EDV] in the trained heart.
More blood enters the left ventricle, and a greater percentage of what enters
is forced out with each concentration, resulting in an increase in stroke
volume.
Adaptatioins in stroke volume during endurance
training are illustrated by a study in which older men endurance trained for
one year. Their cardiovascular function was evaluated before and after
training. The subjects performed running, treadmill, and cycle ergometer
exercise for 1h each day, four days per week. They exercised at intensities of
60% to 80% pf VO2max, with brief bouts of exercise exceeding 90% of
VO2max. End-diastolic volume increased at rest and throughout
submaximal exercise. The ejection fraction increased, which was associated with
a decreased end-systolic volume, suggesting increased contractility of the left
ventricle. VO2max increased by 23%, indicating a substantial
improvement in endurance.
It is clear that central stroke volume
adaptations occur with endurance training, but there are also peripheral
adaptations that contribute to the increase in VO2max, at least in
middle-aged exercisers. This was demonstrated in a unique longitudinal study
involving both exercise training and a bed rest deconditioning model. Five
20-year-old men were tested(baseline values), placed on bed rest for 20
days(deconditioning), and then trained for 60 days, starting immediately at the
conclusion of bed rest. These same five men were restudied 30 years later at
the age of 50; they were tested at baseline in a relatively sedentary state and
after six months of endurance training. The average percentage increases in VO2max
were similar for the subjects at age 20(18%) and at age 50(14%). However, the
increase in VO2max at age 20 was explained by increases in both maximal cardiac
output and maximal(a-ṽ)O2 difference; at age 50, the increase was explained
primarily by an increase in(a-ṽ)O2 difference, while maximal cardiac output was
unchanged. Maximal stroke volume was increased after training at both age 20
and age 50 but to a lesser degree at age 50(+16ml/beat at age 20 vs. +8ml/beat
at age 50).