



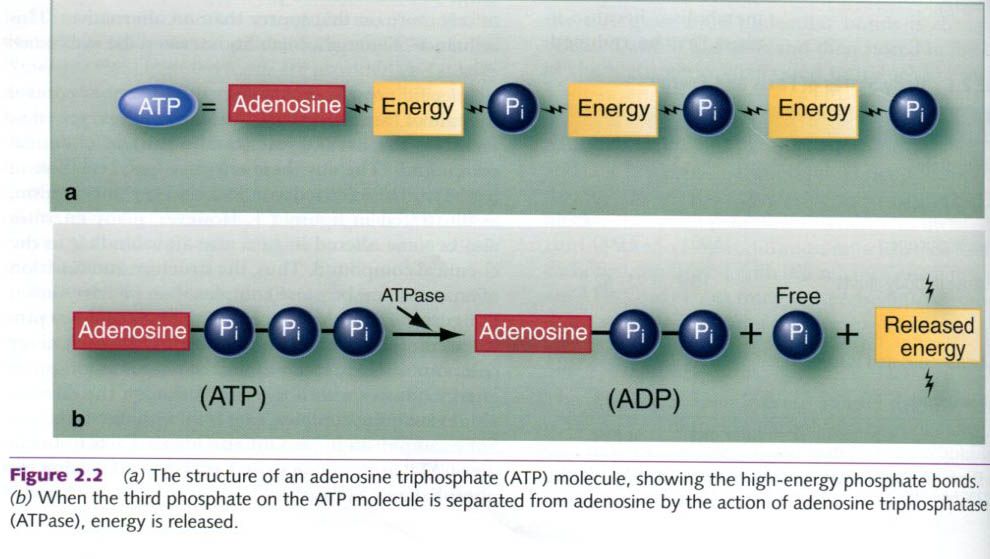
To generate ATP, a phosphate group is added to
the relatively low-energy compound, ADP, a process called phosphorylation. Some ATP is generated independent of oxygen
availability, and such metabolism is called substrate-level phosphorylation. Other
ATP-producing reactions occur without oxygen, a process called anaerobic metabolism. When these
reactions occur with the aid of oxygen, the overall process is called aerobic metabolism, and the aerobic
conversion of ADP to ATP is oxidative
phosphorylation.
Cells generate ATP through three different
processes or systems:
- The ATP-PCr system
- The glycolytic system(glycolysis)
- The oxidative
system(oxidative phosphorylation)
ATP-PCr system
The simplest of the energy systems is the ATP-PCr system, shown in the
picture below. In addition to storing a very small amount of ATP directly,
cells contain another high-energy phosphate molecule that stores energy. This
molecule is called phosphocreatine,
or PCr(sometimes called
creatine phosphate). Unlike freely available ATP, energy released by the
breakdown of PCr is not directly used for cellular work. Instead, it regenerates
ATP to maintain a relatively constant supply.
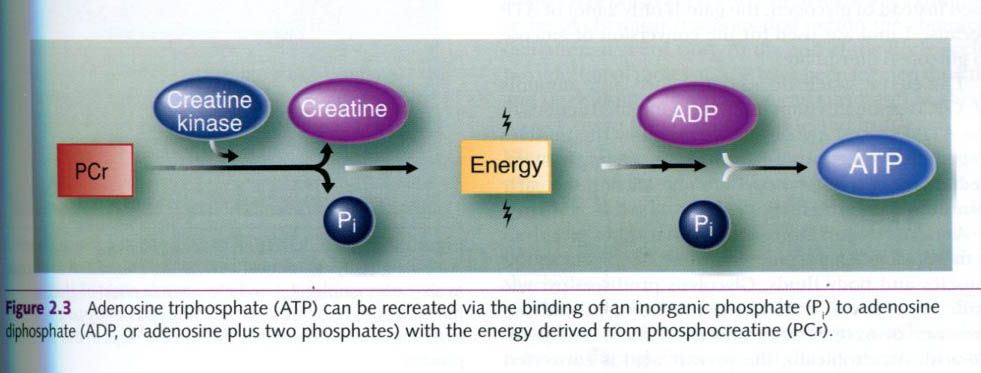
The release of energy from PCr is facilitated
by the enzyme creatine kinase, which
acts on PCr to separate Pi from
creatine. The energy released can then be used to add a Pi molecule
to and ADP molecule, forming ATP. As energy is released from ATP by the
splitting of a phosphate group, cells can prevent ATP depletion by breaking
down PCr, providing energy and Pi to re-form ATP from ADP.
This
process is rapid and can be accomplished without any special structures within
the cell. The ATP-PCr system is classified as substrate-level metabolism.
Although it can occur in the presence of oxygen, this process does not require
oxygen.
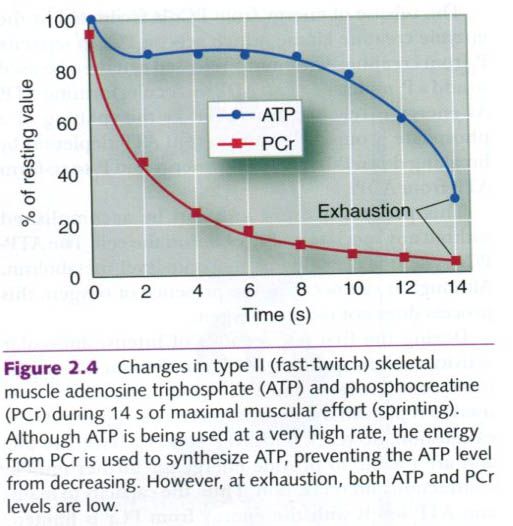
During the first few seconds of intense
muscular activity, such as sprinting, ATP is maintained at a relatively
constant level, but PCr declines steadily as it is used to replenish the
depleted ATP. At exhaustion, however, both
ATP and PCr levels are low and are unable to provide energy for further muscle
contraction and relaxation. Thus, the capacity to maintain ATP levels with the
energy from PCr is limited. The combination of ATP and PCr stores can
sustain the muscles’ energy needs for only 3 to 15s during an all-out sprint.
Beyond that time, muscles must rely on other processes for ATP
formation:glycolitic and oxidative combustion of fuels.
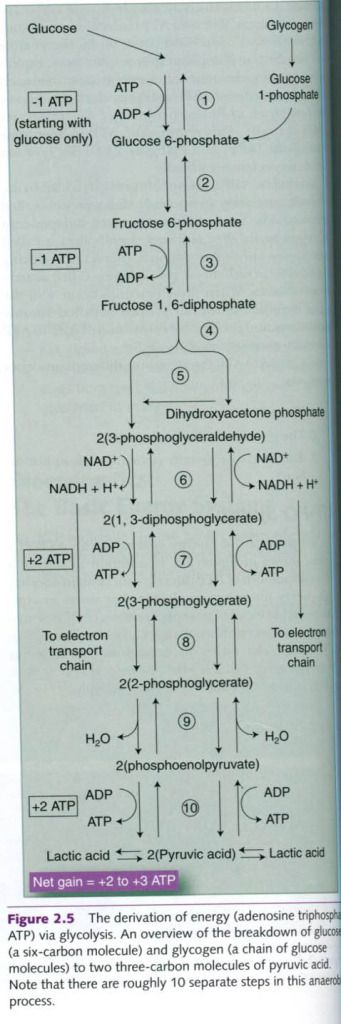
Glycolitic system
Another method of ATP production involves the
liberation of energy through the breakdown(lysis) of glucose. This system is called the glycolitic
system because it entails glycolisis,
which is the breakdown of glucose through a pathway that involves a sequence of
glycolitic enzymes. An overview is possible on the photo below.
Glucose accounts for about 99% of all sugars
circulating in the blood. Blood glucose comes from the digestion of
carbohydrate and the breakdown of liver glycogen. Glycogen is synthesized from
glucose by a process called glycogenolysis. Glycogen is stored in the liver or
in muscle until needed. At that time, the glycogen is broken down to
glucose-I-phosphate, which enters the glycolysis pathway, a process termed glycogenolysis.
Before either glucose or glycogen can be used
to generate energy, it must be converted to a compound called
glucose-6-phosphate. Even though the goal of glycolysis is to release ATP, the
conversion of a molecule of glucose to glucose-6-phosphate requires one ATP
molecule. In the conversion of glycogen, glucose-6-phosphate is formed from
glucose-1-phosphate without this energy expenditure. Glycolysis technically
begins once the glucose-6-phosphate is formed.
Glycolysis, which is far more complex than the
ATP-PCr system, requires 10-12 enzymatic reactions for the breakdown of
glycogen to lactic acid. All these enzymes operate within the cell cytoplasm.
The net gain from this process is 3 moles(mol) of ATP formed for each mole of
glycogen broken down. If glucose is used instead of glycogen, the gain is only
2 mol of ATP because 1 mol was used for the conversion of glucose to
glucose-6-phosphate.
This energy system does not produce large
amounts of ATP. Despite this limitation, the combined actions of the ATP-PCr
and glycolytic systems allow the muscles to generate force even when the oxygen
supply is limited. These two systems predominate during the early minutes of
high-intensity exercise.
Another major limitation of anaerobic
glycolysis is that it causes an accumulation of lactic acid in the muscles and
body fluids. Glycolysis produces pyruvic acid. This process does not require
oxygen, but the presence of oxygen determines the fate of the pyruvic acid.
Anaerobically, the pyruvic acid is converted directly to lactic acid, an acid
with the chemical formula C3H6O3. When lactic
acid releases hydrogen ions(Na+) or potassium ions(K+) to
form a salt, called lactate. Anaerobic glycolysis produces lactic acid, but it
quickly dissociates, and lactate is formed. For this reason, the terms often
are used interchangeably, although they do not refer to the same molecule.
In all-out sprint events lasting 1 or 2 min,
the demands on the glycolytic system are high, and muscle lactic acid
concentrations can increase from a resting value of about 1 mmol/kg of muscle
to more than 25 mmol/kg. This acidification of muscle fibers inhibits further
glycogen breakdown because it impairs glycolytic enzyme function. In addition,
the acid decreases the fibers’ calcium-binding capacity and thus may impede
muscle contraction.
A muscle fiber’s rate of energy use during
exercise can be 200 times greater than at rest. The ATP-PCr and glycolitic systems
alone cannot supply all the needed energy. Furthermore, these two systems are
not capable of supplying all of the energy needs for all-out activity lasting
more than 2 min or so. Prolonged exercise relies on the third energy system,
the oxidative system.
Oxidative system
The final system of cellular energy production
is the oxidative system. This is
the most complex of the three energy systems, and only a brief overview is
provided here. The process by which the body breaks down substrates with the
aid of oxygen to generate energy is called cellular
respiration. Because oxygen is used, this is an aerobic process. This
oxidative production of ATP occurs within special cell organelles called
mitochondria. In muscles, these are adjacent to the myofibrils and are also
scattered throughout the sarcoplasm.
Muscle need a steady supply of energy to
continuously produce the force needed during long-term activity. Unlike
anaerobic ATP production, the oxidative system is slow to turn on; but it has a
tremendous energy-yielding capacity, so aerobic metabolism is the primary
method of energy production during endurance events. This places
considerable demands on the cardiovascular and respiratory systems to deliver
oxygen to the active muscles.
Oxidation of carbohydrate
Oxidative production of ATP involves three
processes:
- Aerobic glycolysis
- The Krebs cycle
- The electron transport chain
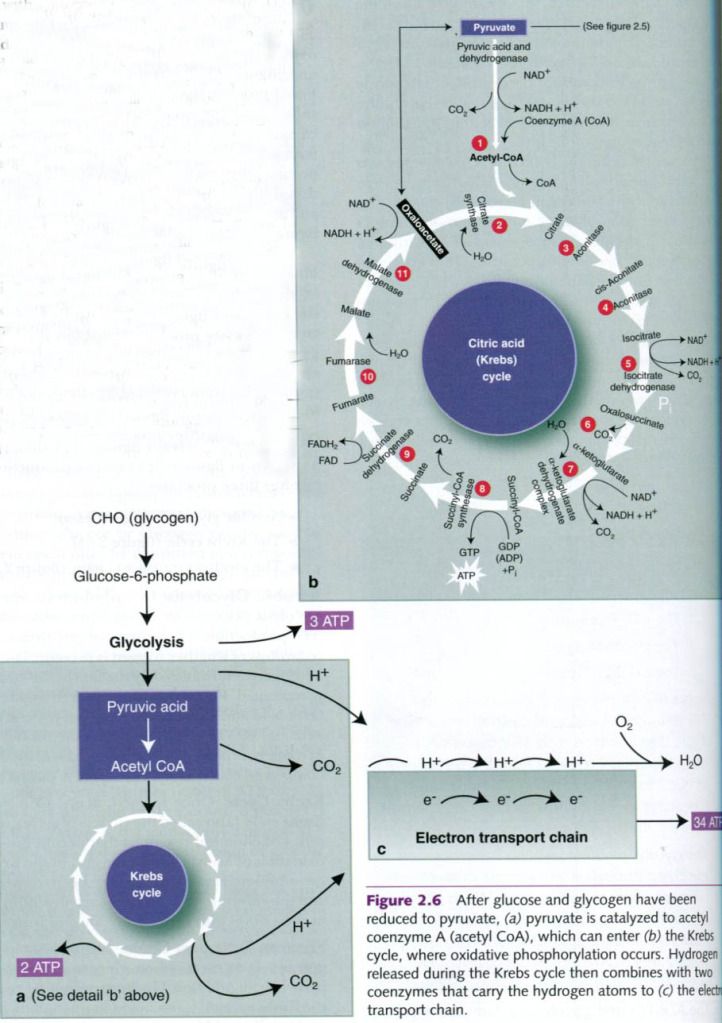
Aerobic
glycolysis – In
carbohydrate metabolism, glycolysis plays a role in both anaerobic and aerobic
ATP production. The process of glycolysis is the same regardless of whether
oxygen is present. The presence of oxygen determines only the fate of the end
product, pyruvic acid. Recall that anaerobic glycolysis produces lactic acid
and only 3 mol of ATP per mole of glycogen, or 2 mol of ATP per mole of
glucose. In the presence of oxygen, however, the pyruvic acid is converted into
a compound called acetyl coenzyme
A(acetyl CoA).
Krebs
Cycle – Once formed,
acetyl CoA enters the Krebs cycle(also
called citric acid cycle), a complex series of chemical reactions that
permit the complete oxidation of acetyl CoA. At the end of the Krebs
cycle, two additional moles of ATP have been formed recently, and the
substrate(the original carbohydrate) has been broken down into carbon dioxide
and hydrogen.
Electron
Transport Chain – During
glycolysis, hydrogen ion is released when glucose is metabolized to pyruvic
acid. Additional hydrogen ion is released during the Krebs cycle. If it
remained in the system, the inside of the cell would become too acidic. What
happens to this hydrogen?
The Krebs cycle is coupled to a series of
reactions known as the electron
transport chain. The hydrogen released during glycolysis and during the
Krebs cycle combines with two coenzymes: nicotinamide adenine dinucleotide(NAD) and flavin adenine
dinucleotide(FAD). These carry the hydrogen atoms to the electron
transport chain, where they are split into protons and electrons. At the end of
the chain, the H+ combines with oxygen to form water, thus
preventing acidification. The electrons that were split from the hydrogen pass
through a chain of reactions(hence the name electron transport chain) and
ultimately provide energy for the phosphorylatioon of ADP, thus forming ATP.
Because this process relies on oxygen, it is reffered to as oxidative phosphorylation.

Energy
Yield From Oxydation of Carbohydrate – The complete oxidation of carbohydrate can generate 37 to 39
molecules of ATP from one molecule of muscle glycogen. If the process
begins with glucose, the maximal net gain is 38 ATP molecules(recall that
one ATP molecule is used for conversion to glucose-6-phosphate before
glycolysis begins).
It should be noted that the molecules of
reduced NAD(termed NADH) formed in the cytoplasm cannot directly enter the
mitochondria. They must donate their electrons to either NADH or reduced
FAD(FADH) carrier molecules in the electron transport chain. Two cytoplasmic NADH yield six ATP
molecules, as opposed to only four ATP molecules when their electrons are
donated to mitochondrial FADH. Thus, when FADH is carrier, only up to 36 ATP molecules can be
generated from glucose and 37 ATP molecules from glycogen.
Energy
production from the oxidation of muscle glycogen
|
||
Stage
of process
|
Direct
|
By
oxidative phosphorylation
|
Glycolysis(glucose
to pyruvic acid)
|
3
|
4-6b
|
Pyruvic
acid to acetyl coenzyme A
|
0
|
6
|
Krebs
cycle
|
2
|
22
|
Subtotal
|
5
|
32-34
|
Total
|
37 -39
|
|
Oxidation of fat
As noted earlier, fat also contributes
importantly to muscles’ energy needs.
Muscle and liver glycogen stores can provide only approximately 2,500 kcal of
energy, but the fat stored inside muscle fibers and in fat cells can supply at
least 70,000 to 75,000 kcal, even in a lean adult.
Although many chemical compounds(such as
triglycerides, phospholipids, and cholesterol) are classified as fats, only
triglycerides are major energy sources. Triglycerides
are stored in fat cells and between and within skeletal muscle fibers. To
be used for energy, a triglyceride must be broken down to its basic units; one
molecule of glycerol and three FFA molecules. This process is called lypolysis, and it is carried out by
enzymes known as lipases.
Free
fatty acids are the primary
energy source. Once liberated from glycerol, FFAs can enter the blood
and be transported throughout the body, entering muscle fibers by simple
diffusion or by transporter-mediated(facilitated) diffusion. Their rate of
entry into the muscle fibers depends on the concentration gradient. Increasing
the concentration of FFAs in the blood increases the rate of their transport
into muscle fibers.
Β – Oxidation – Although the various FFAs in the body differ structurally, their
metabolism is essentially the same, as shown in the left half of figure below.
On entering the muscle fiber, FFAs are enzymatically activated with energy from
ATP, preparing them for catabolism(breakdown) within the mitochondria. This
enzymatic catabolism of fat by the mitochondria is termed Β–oxidation.
In this process, the carbon
chain of an FFA is cleaved into separate two-carbon units of acetic acid. For
example, if an FFA originally has a 16-carbon chain, Β–oxidation yields eight
molecules to acetyl CoA.
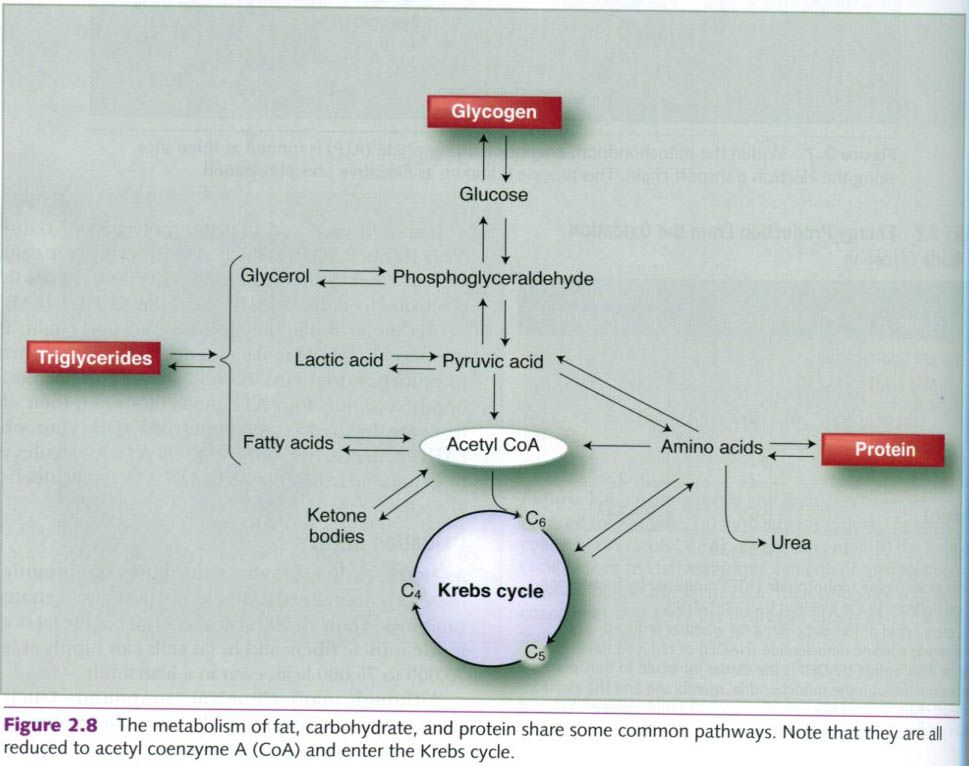
Krebs Cycle and the Electron Transport Chain – From this point on, fat
metabolism follows the same path as oxidative carbohydrate metabolism. Acetyl CoA formed by Β–oxidation enters the Krebs cycle. The Krebs cycle generates hydrogen,
which is transported to the electron transport chain along with the hydrogen
generated during Β–oxidation to undergo oxidative phosphorylation. As in glucose
metabolism, the by-products of FFA oxidation are ATP, H2O, and
carbon dioxide(CO2). However, the complete combustion of an FFA
molecule requires more oxygen because an FFA molecule contains considerably
more carbon than a glucose molecule.
The advantage of having more
carbon in FFAs than in glucose is that more acetyl CoA is formed from the
metabolism of a given amount of fat, so more acetyl CoA enters the Krebs cycle
and more electrons are sent to the electron transport chain. This is why fat
metabolism can generate so much more energy than glucose metabolism. Unlike
glycogen, fast are heterogeneous, and the amount of ATP produced depends on the
specific fat oxidized.
Consider the example of palmitic
acid, a rather abundant 16-carbon FFA. The
combined reactions of oxidation, the Krebs cycle, and the electron transport
chain produce 129 molecules of ATP from one molecule of palmitic acid(shown in
the table below), compared with only 38 molecules of ATP from glucose or 39
from glycogen.
Energy
production from the oxidation of palmitic acid
|
||
Adenosine
triphosphate produced from one molecule of C16H32O2
|
||
Stage
of process
|
Direct
|
By
oxidative phosphorylation
|
Fatty
acid activation
|
0
|
-2
|
Β–oxidation
|
0
|
35
|
Krebs
cycle
|
8
|
88
|
Subtotal
|
8
|
121
|
Total
|
129
|
|
Oxidation of protein
As noted earlier, carbohydrates and fatty acids
are the preferred fuels. But proteins, or rather the amino acids that compose
proteins, are also used. Some amino acids can be converted into glucose(by
gluconeogenesis). Alternatively, some can be converted into various intermediates
of oxidative metabolism(such as pyruvate or acetyl CoA) to enter the oxidative
process.
Protein’s
energy yield is not as easily determined as that of carbohydrate or fat because
protein also contains nitrogen.
When amino acids are catabolized, some of the released nitrogen is used to
form new amino acids, but the remaining nitrogen cannot be oxidized by the
body. Instead it is converted into urea and then excreted, primarily in the
urine. This conversion requires the use of ATP, so some energy is spent in this
process.
When protein is broken down through combustion
in the laboratory, the energy yield is 5.65 kcal/g. However, because of the
energy expended in converting nitrogen to urea, when protein is metabolized in
the body, the energy yield is only about 4.1 kcal/g, 27.4% less than the
laboratory value.
To accurately assess the rate of protein
metabolism, the amount of nitrogen being eliminated from the body must be
determined. These measurements require urine collection for 12 to 24h periods,
a time-consumption process. Because the healthy body uses little protein during
rest and exercise(usually not more than 5% of total energy expended), estimates
of total energy expenditure generally ignore protein metabolism.
Interaction of the three energy systems
The three energy systems do not work
independently of one another. When a person is exercising at the highest
intensity possible, from the shortest sprints(less than 10s) to endurance
events(greater than 30 min), each of the energy systems is contributing to the
total energy needs of the body. Generally one energy system dominates, except
when there is a transition from the predominance of one energy system to
another. As an example, in a 10s, 100m sprint, the ATP-PCr system is the
predominant energy system, but both the anaerobic glycolytic and oxidative
systems provide a small portion of the energy needed. At the other extreme, in
a 30 min, 10,000m (10,936 yd) run, the the oxidative system is predominant, but
both the ATP-PCr and anaerobic glycolytic systems contribute some energy as
well.
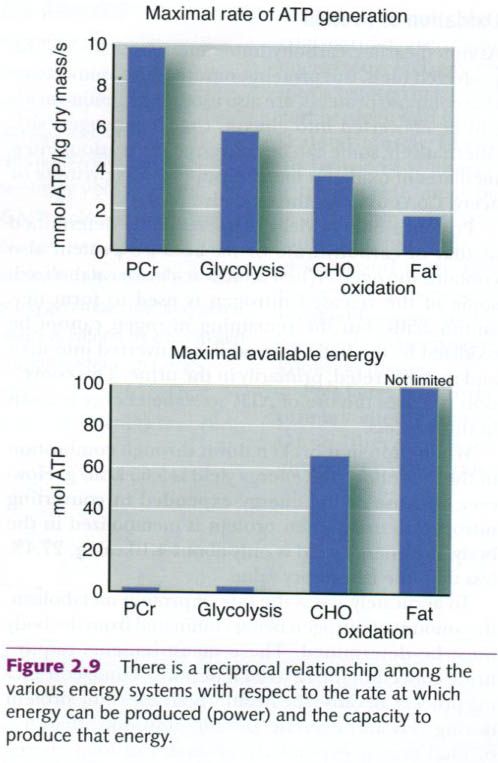
Figure below shows reciprocal relationship
among the energy systems with respect to power and capacity. The PCR energy
system can provide energy at a fast rate but has a low capacity for energy
production. Thus it supports exercise that is intense but of very short
duration. By contrast, fat oxidation takes longer to gear up and produces
energy at a slower rate; however, the amount of energy it can produce is
unlimited.
Oxidative capacity of muscle
We have seen that the processes of oxidative
metabolism have the highest energy yields. It would be ideal if these processes
always functioned at peak capacity. But, as with all physiological systems,
they operate within certain constraints. The oxidative capacity of muscle(QO2)
is a measure of its maximal capacity to use oxygen. This measurement is made in
the laboratory where a small amount of muscle tissue can be tested to determine
its capacity to consume oxygen when chemically stimulated to generate ATP.
Enzyme activity
Muscle fibers’ capacity to oxidize carbohydrate
and fat is difficult to determine. Numerous studies have shown a close
relationship between a muscle’s ability to perform prolonged aerobic exercise
and the activity of its oxidative enzymes. Because many enzymes are required
for oxidation, the enzyme activity of the muscle fibers provides a reasonable
indication of their oxidative potential.
Measuring all the enzymes in muscles is
impossible, so a few representative enzymes have been selected to reflect the
aerobic capacity of the fibers. The enzymes most frequently measured include
succinate dehydrogenase and citrate synthase, mitochondrial enzymes involved in
the Krebs cycle. This picture below illustrates the close relationship between
succinate dehydrogenase activity in the vastus lateralis muscle and the
muscle’s oxidative capacity. Endurance athletes’ muscles have oxidative enzyme
activities nearly two to four times greater than those of untrained men and
women.

Fiber type composition and endurance training
A muscle’s fiber type composition primarily
determines its oxidative capacity. As noted in muscle fiber thread,
slow-twitch, or type I, fibers have a greater capacity for aerobic activity
than the fast-twitch, or type II, fibers because type I fibers have more
mitochondria and higher concentrations of oxidative enzymes. Type II fibers are
better suited for glycolitic energy production. Thus, in general, the more type
I fibers in one’s muscles, the greater the oxidative capacity of those muscles.
Elite distance runners,for example, have been reported to possess more type I
fibers, more mitochondria, and higher muscle oxidative enzyme activities than
do untrained individuals.
Endurance training enhances the oxidative capacity
of all fibers, especially type II fibers. Training that places demands on
oxidative phosphorylation stimulates the muscle fibers to develop more
mitochondria that also are larger and contain more oxidative enzymes. By
increasing the fibers’ enzymes for Β–oxidation , this training also
enables the muscle to rely more heavily on fat for ATP production. Thus, with
endurance training, even people with large percentages of type II fibers can
increase their muscles’ aerobic capacities. But it is generally agreed that an
endurance-trained type II fiber will not develop the same high endurance
capacity as a similarly trained type I fiber.
Oxygen needs
Although the oxidative capacity
of a muscle is determined by the number of mitochondria and the amount of oxidative
enzymes present, oxidative metabolism ultimately depends on an adequate supply
for oxygen. At rest, the need for ATP is relatively small, requiring minimal
oxygen delivery. As exercise intensity increases, so do energy demands. To meet
them, the rate of oxidative ATP production increases. In an effort to “meet the
muscles” need for oxygen, the rate and depth of respiration increase, improving
gas exchange in the lungs, and the heart beats faster and more forcefully,
pumping more oxygenated blood to the muscles. Arterioles dilate to facilitate
delivery of arterial blood to muscle capillaries.
The human body stores little
oxygen. Therefore, the amount of oxygen entering the blood as it passes through
the lungs is directly proportional to the amount used by the tissues for
oxidative metabolism. Consequently, a reasonably accurate estimate of aerobic
energy production can be made by measuring the amount of oxygen consumed at the
lungs.
“Physiology of sport and exercise”, fourth
edition; Jack H. Wilmore, David L. Costill, W. Larry Kenney






0 коментара:
Постави коментар