



Ask most exercisers what causes fatique during
exercise, and the most common answer involves two words: lactic acid. Not only
is this common misconception an oversimplification, but there is mounting
evidence that lactic acid may actually have beneficial effects on exercise
performance!
Fatique is an extremely complex phenomenon.
Most efforts to describe underlying causes and sites of fatique have focused
on:
- Energy delivery( ATP-PCr,
anaerobic glycolysis, and oxidation);
- Accumulation of metabolic
by-products, such as lactate and H+;
- Failure of the muscle fiber’s
contractile mechanism;
- Alterations in the nervous
system.
The first three causes occur within the muscle
itself and are often referred to as peripheral fatique. Changes in the nervous
system may cause central fatique. None of these alone can explain all aspects
of fatique, and several causes may act synergistically to cause fatique.
Fatique is rarely caused by a single factor but typically by multiple factors
acting at multiple sites. Mechanisms of fatique depend on the type and
intensity of the exercise, the fiber type of the involved muscles, the
subject’s training status, and even his or her diet. Many questions about
fatique remain unanswered, especially about cellular sites of fatique within
the muscle fibers themselves.
Energy systems and fatique
The energy systems are an obvious area to
explore when one is considering possible causes of fatique. When we are well
fatiqued, we often express this by saying:” I have no energy.” But this use of
term energy is far removed from its psychological meaning.
PCr depletion
Recall that PCr depletion is used under
anaerobic conditions, such as short-term high-intensity effort, to rebuild ATP
as it is used and thus maintain ATP stores within the muscle. Biopsy studies of
human thigh muscles have shown that during repeated maximal contractions,
fatique coincidences with PCr depletion. Although ATP is directly responsible
for the energy used during such activities, it is depleted less rapidly than
PCr during muscular effort because ATP is being produced by other systems. But
as PCr is depleted, the ability to quickly replace the spent ATP is seriously
hindered. Use of ATP continues, but the ATP-PCr system is less able to replace
it. Thus, ATP levels also decrease. At exhaustion, both ATP and PCr may be
depleted. It now appears that Pi, which increases during intense short-term
exercise because of the breakdown of PCr, is a potential cause of fatique in
this type of exercise.
To delay fatique, the athlete must control the
rate of effort through proper pacing to ensure that PCr and ATP are not
prematurely exhausted. This holds true even in endurance-type events. If the
beginning pace is too rapid, available ATP and PCr concentrations will quickly
decrease, leading to early fatique and an inability to maintain the pace in the
event’s final stages. Training and experience allow the athlete to judge the
optimal pace that permits the most efficient use of ATP and PCr for the entire
event.
Glycogen depletion
Muscle ATP concentrations are also maintained
by the aerobic and anaerobic breakdown of muscle glycogen. In events lasting
longer than a few seconds, muscle glycogen becomes the primary energy source
for ATP synthesis. Unfortunately, glycogen reserves are limited and are
depleted quickly. Since the muscle biopsy technique was first established,
studies have shown a correlation between muscle glycogen depletion and fatique
during prolonged exercise.
As with PCr use, the rate of muscle glycogen
depletion is controlled by the intensity of the activity. Increasing the
intensity results in a disproportionate decrease in muscle glycogen. During
sprint running, for example, muscle glycogen may be used 35 to 40 times faster
than during walking. Muscle glycogen can be a limiting factor even during mild
effort. The muscle depends on a constant supply of glycogen to meet the high
energy demands of exercise.
Muscle glycogen is used more rapidly during the
first few minutes of exercise than in the later stages, as seen in figure
below. The illustration shows the change in muscle glycogen content in the
subject’s gastrocnemius(calf) muscle during the test. Although the subject ran
the test at a steady pace, the rate of muscle glycogen metabolized from the
gastrocnemius was greatest during the first 75 min.
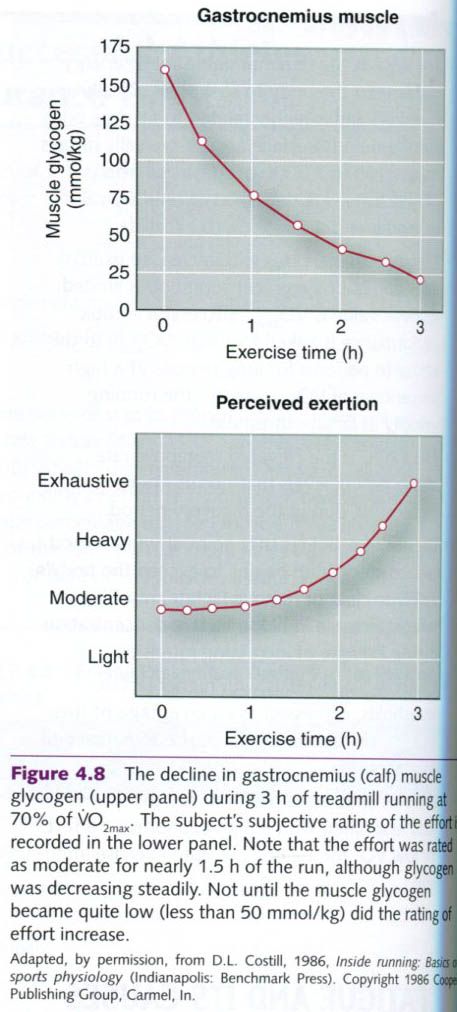
The subject also reported his perceived
exertion(how difficult his effort seemed to be) at various times during the
test. He felt only moderately stressed early in the run, when his glycogen
stores were still high, even though he was using glycogen at a high rate. He
did not perceive severe fatique until his muscle glycogen levels were nearly
depleted. Thus, the sensation of fatique in long-term exercise conincides with
a decreased concentration of muscle glycogen, but not with its rate of
depletion. Marathon runners commonly refer to
the sudden onset of fatique that they experience at 29 to 35 km(18-22 mi) as
“hitting the wall”. At least part of this sensation can be attributed to muscle
glycogen depletion.
Glycogen
depletion in different fiber types
Muscle fibers are recruited and deplete their
energy reserves in selected patterns. The individual fibers most frequently
recruited during exercise may become depleted of glycogen. This reduces the
number of fibers capable of producing the muscular force needed for exercise.
This glycogen depletion is illustrated in the
figure below, which shows a micrograph of muscle fibers taken from a runner
after a 30km(18.6 miles) run. Figure a has been stained to differentiate type I
and type II fibers. One of the type II fibers is circled. Figure b shows a
second sample from the same muscle, stained to show glycogen. The
redder(darker) the stain, the more glycogen is present. Before the run, all
fibers were full of glycogen and appeared red(not depicted). In figure b(after
the run), the lighter type I fibers are almost completely depleted of glycogen.
This suggests that type I fibers are used more heavily during endurance
exercise that requires only moderate force development, such as the 30km run.
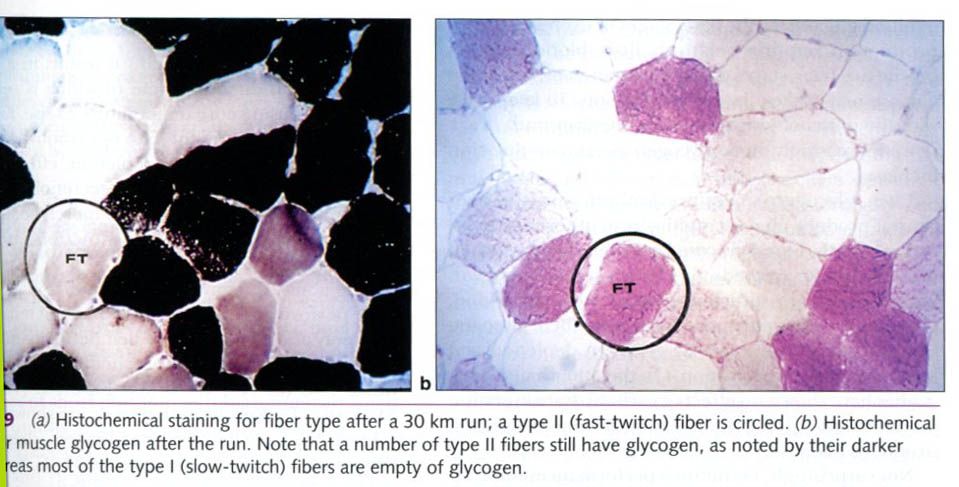
The pattern of glycogen depletion from type I
and type II fibers depends on the exercise intensity. Recall that type I fibers
are the first fibers to be recruited during light exercise. As muscle tension
requirements increase, type IIa fibers are added to the workforce. In exercise
approaching maximal intensities, the type IIx fibers are added to the pool of
recruited fibers.
Depletion
in different muscle groups
In addition to selectively depleting glycogen from type I or type II fibers,
exercise may place unusually heavy demands on select muscle groups. In one
study, subjects ran on a treadmill positioned for uphill, downhill, and level
running for 2h at 70% of VO2max. Figure below compares the resultant
glycogen depletion in three muscles of the lower extremity: the vastus
lateralis(knee extensor), the gastrocnemius(ankle extensor), and the
soleus(another knee extensor).

The results show that whether one runs uphill,
downhill, or on a level surface, the gastrocnemius uses more glycogen than does
the vastus lateralis or the soleus. This suggests that the ankle extensor
muscles are more likely to become depleted during distance running than are the
thigh muscles, isolating the site of fatique to the lower leg muscles.
Glycogen
depletion and blood glucose
Muscle glycogen alone cannot provide enough carbohydrate for exercise lasting
several hours. Glucose delivered by the blood to the muscles contributes a lot
of energy during endurance exercise. The liver breaks down its stored glycogen
to provide a constant supply of blood glucose. In the early stages of exercise,
energy production requires relatively little blood glucose; but in later stages
of an endurance event, blood glucose may make a large contribution. To keep
pace with the muscles’ glucose uptake, the liver mus break down increasingly
more glycogen as exercise duration increases.
Liver glycogen stores are limited, and the
liver cannot produce glucose rapidly from other substrates. Consequently, blood
glucose levels can decrease when muscle uptake exceeds the liver’s glucose
output. Unable to obtain sufficient glucose from the blood, the muscles must
rely more heavily on their glycogen reserves, accelerating muscle glycogen depletion and leading to earlier exhaustion. On the
other hand, most studies have shown no effect of carbohydrate ingestion on net muscle glycogen utilization during
prolonged, strenuous exercise.
Not surprisingly, endurance performances
improve when the muscle glycogen supply is elevated before the start of
activity. Glycogen depletion and hypoglycemia(low blood sugar) limit
performance in activities lasting longer than 60 to 90 min.
Mechanisms
of fatique with glycogen depletion
It does not appear likely that glycogen
depletion directly causes fatique during endurance exercise performance.
Rather, the depletion of muscle glycogen
may be the first step in a series of events that leads to fatique. A certain
level of muscle glycogen metabolism is necessary to maintain oxidative
metabolism of both carbohydrates and fats using Krebs cycle. That is, we now know that a certain rate of glycogen
breakdown is needed for the optimal production of reduced nicotinamide adenine
dinucleotide(NADH) and to maintain the electron transport system.
Additionally, as glycogen is depleted,
exercising muscle relies more heavily on the metabolism of FFAs. To accomplish
this, more FFAs must be moved into the mitochondria, and the rate of transfer
may limit FFA oxidation to the point where it can no longer keep up with the
need for fat oxidation.
Metabolic by-products and fatique
Various by-products of metabolism have been
implicated as factors causing, or contributing to, fatique. One example is Pi, which increases
during intense short-term exercise as PCr and ATP are being broken down.
Additional metabolic by-products that have received the most attention in
discussing fatique are heat, lactate and hydrogen ions.
Heat, muscle temperature, and fatique
Recall that energy expenditure results in a
relatively large heat production, some of which is retained in the body,
causing core temperature to rise. Exercise in the heat can increase the rate of
carbohydrate utilization and hasten glycogen depletion, effects that may be
stimulated by the increased secretion of epinephrine. It is hypothesized that
high muscle temperature impair both skeletal muscle function and muscle
metabolism.
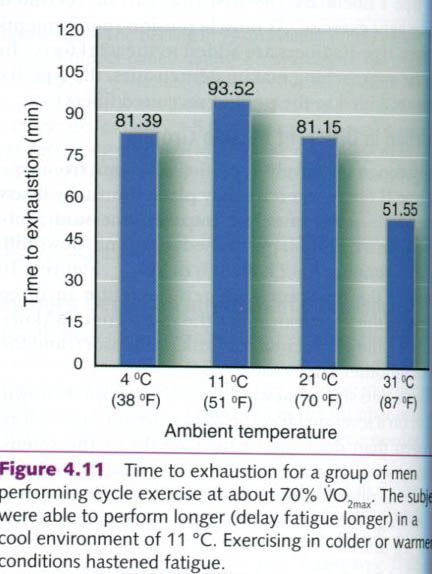
The ability to continue moderate-to
high-intensity cycle performance is affected by ambient temperature. Galloway
and Maughan studied performance time to exhaustion of male cyclists at four different air temperatures: 4°C(38°F),
11°C(51°F), 21°C(70°F), and 31°C(87°F). Results are shown below. Time to
exhaustion was longest when subject exercised on air temperature of 11°C, but
lower at colder and warmer temperatures. Fatique set in earliest at 31°C.
Precooling of muscles similarly prolonged exercise, while preheating causes
earlier fatique. Heat acclimation spares glycogen and reduces lactate
accumulation.
Lactic acid, hydrogen ions and fatique
Recall that lactic acid is a by-product of
anaerobic glycolysis. Although most people believe that lactic acid is
responsible for fatique in all types of exercise, lactic acid accumulates
within the muscle fiber only during relatively brief, highly intense muscular
effort. Marathon runners, for example, may
have near-resting lactic acid levels at the end of the race, despite their
fatique. Their fatique is caused most likely by inadequate energy supply, not lactic acid.
Short sprints in running, cycling, and swimming
all lead to large accumulations of lactic acid. But the presence of lactic acid
should not be blamed for the feeling of fatique in itself. When not cleared,
the lactic acid dissociates, converting to lactate acid causing an accumulation
of hydrogen ions. This H+ accumulation causes muscle acidification,
resulting in a condition known as acidosis.
Activities of short duration and high
intensity, such as sprint running and sprint swimming, depend heavily on
anaerobic glycolysis and produce large amounts of lactate and H+
within the muscles. Fortunately, the cells and body fluids possess buffers,
such as bicarbonate(HCO3), that minimize the disrupting influence of
the H+. Without these buffers, H+ would lower the pH to about 1.5,
killing the cells. Because of the body’s buffering capacity, the H+
concentration remains low even during the most severe exercise, allowing muscle
pH to decrease from a resting value of 7.1 to no lower than 6.6 to 6.4 at
exhaustion.
However, pH changes of this magnitude adversely
affect energy production and muscle contraction. An intracellular pH below 6.9
inhibits the action of phosphofructokinase, an important glycolytic enzyme,
slowing the rate of glycolysis and ATP production. At a pH of 6.4, the
influence of H+ stops any further glycogen breakdown, causing a
rapid decrease in ATP and ultimately exhaustion. In addition, H+ may displace calcium within the fiber, interfering
with the coupling of the actin-myosin cross-bridges and decreasing the muscle’s
contractile force. Most researchers agree that low muscle pH is the major
limiter of performance and the primary cause of fatique during maximal, all-out
exercise lasting more than 20s to 30s.
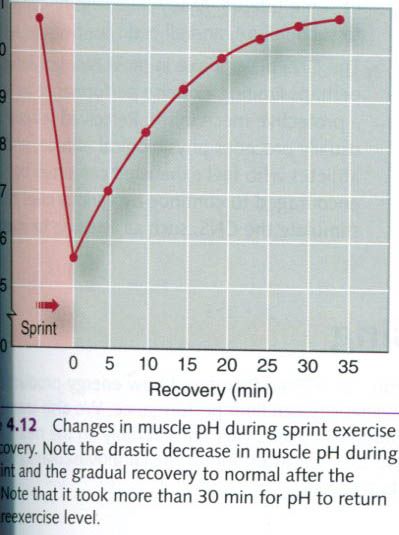
As seen in the figure above, reestablishing the
preexercise muscle pH after an exhaustive sprint bout requires about 30 to 35
min of recovery. Even when normal pH is restored, blood and muscle lactate
levels can remain quite elevated. However, experience has shown that an athlete
can continue to exercise at relatively high intensities even with a muscle pH
below 7.0, and a blood lactate level above 6 or 7 mmol, four to five times the
resting value.
Some coaches and sport physiologists have
attempted to use blood lactate measurements to gauge the intensity and volume
of training needed to produce an optimal training stimulus. Such measurements
provide an index of training intensity, but they might not reflect the
anaerobic processes of the state of acidosis in the muscles. Because lactate
and H+ are generated in the muscles, both diffuse in the body fluids
and transported to other areas of the body to be metabolized. Consequently,
blood lactate concentrations depend on the rates of production, diffusion,
oxidation, and clearance. A variety of factors can influence these processes,
so measuring blood lactate is of questionable value for fine-tuning training.
Neuromuscular fatique
Thus we have considered only factors within the
muscle that might be responsible for fatique. Evidence also suggests that under
some circumstances, fatique may result from an inability to activate the muscle
fibers, a function of the nervous system. The neural impulse is transmitted
across the neuromuscular junction to activate the fiber’s membrane, and it
causes the fiber’s sarcoplasmic reticulum to release calcium. The calcium, in turn, binds with troponin to initiate
muscle contraction. Two of several possible neural mechanisms that could
disrupt this process and possibly contribute to fatique are described in
following part.
Neural transmission
Fatique may occur at the neuromuscular
junction, preventing nerve impulse transmission to the muscle fiber membrane.
Studies in the early 1900s clearly established such a failure of nerve impulse
transmission in fatiqued muscle. This failure may involve one or more of the
following processes:
- The release of synthesis of
acetylholine(ACh), the neurotransmitter that relays the nerve impulse from
the motor nerve to the muscle membrane, might be reduced;
- Cholinesterase, the enzyme
that breaks down ACh once it has relayed the impulse, might become
hyperactive, preventing sufficient concentration of ACh to initiate an action potential;
- Cholinesterase activity
might become hypoactive(inhibited), allowing ACh to accumulate
excessively, inhibiting relaxation;
- The muscle fiber membrane
might develop a higher threshold for stimulation by motor neurons;
- Some substance might compete
with ACh for the receptors on the muscle membrane without activating the
membrane;
- Potassium might leave the intracellular space of
the contracting muscle, decreasing the membrane potential to half of its
resting value.
Although most of these causes for a
neuromuscular block have been associated with neuromuscular diseases(such as
myasthenia gravis), they may also cause some forms of neuromuscular fatique.
Some evidence suggests that fatique also may be attributable to calcium retention
within the sarcoplasmic reticulum, which would decrease the calcium available for muscle
contraction. In fact, depletion of PCr and lactate buildup might simply
increase the rate of calcium accumulation within the sarcoplasmic reticulum.
However, these theories of fatique
remain speculative.
Central nervous system
The central nervous system(CNS) also might be a
site of fatique. Undoubtedly, there is some CNS involvement in most types of
fatique. When a subject’s muscles appear to be nearly exhausted, verbal
encouragement, shouting, playing of music, or even direct electrical
stimulation of the muscle can increase the strength of muscle contraction. The
precise mechanisms underlying the CNS role in causing, sensing and even
overriding fatique are not fully understood.
The recruitment of muscle depends, in part, on
conscious control. The stress of exhaustive exercise may lead to conscious or
subconscious inhibition of the athlete’s willingness to tolerate further pain.
The CNS may slow the exercise pace to a tolerable level to protect the athlete.
Indeed, researchers generally agree that the perceived discomfort of fatique
precedes the onset of a physiological limitation within the muscles. Unless
they are highly motivated, most individuals terminate exercise before their
muscles are physiologically exhausted. To achieve peak performance, athletes
train to learn proper pacing and tolerance for fatique.








0 коментара:
Постави коментар