



Oxygen is transported by the blood either
combined with hemoglobin in the red blood cells(greater than 98%) or dissolved
in the blood plasma(2%). Only about 3ml of oxygen are dissolved in each liter
of plasma. Assuming a total plasma volume of 3 to 5L, only about 9 to 15ml of
oxygen can be carried in the dissolved state. This limited amount of oxygen
cannot adequately meet the needs of even resting body tissues, which generally
require more than 250ml of oxygen per minute(depending on body size). However, hemoglobin,
a protein contained within each of the body’s 4 to 6 billion red blood cells,
allows the blood to transport nearly 70 times more oxygen than can be dissolved
in plasma.
Hemoglobin saturation
As just noted, over 98% of oxygen is
transported in the blood bound to hemoglobin. Each molecule of hemoglobin can
carry four molecules of oxygen. When oxygen binds to hemoglobin, it forms
oxyhemoglobin; hemoglobin that is not bound to oxygen is reffered to as
deoxyhemoglobin. The binding of oxygen to hemoglobin depends on the PO2
in the blood and the bonding strength, or affinity, between hemoglobin and
oxygen. The curve in figure below is and oxygen-hemoglobin dissociation curve,
which shows the amount of hemoglobin saturated with oxygen at different PO2
values. The shape of the curve is extremely important for its function in the
body. The relatively flat upper position means that, at high PO2s
such as in the lungs, large drops in PO2 result in only small
changes in hemoglobin saturation. This is called the “loading” portion of the
curve. A high blood PO2 results in almost complete hemoglobin
saturation, which means that the maximal amount of oxygen is bound. But as the
PO2 decreases, so does hemoglobin saturation.
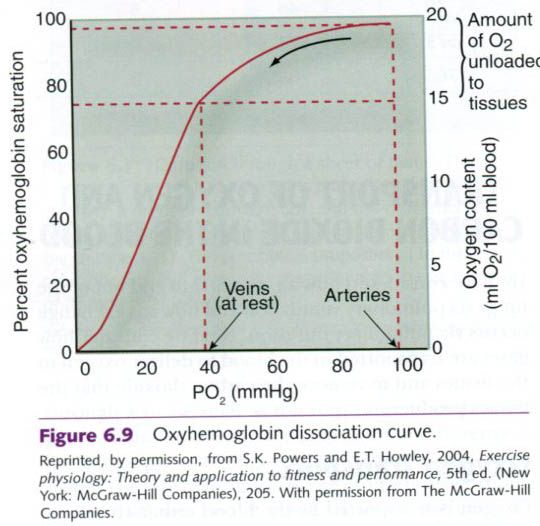
The steep portion of the curve coincides with
PO2 values typically found in the tissues of the body. Here,
relatively small changes in PO2 result in large changes in
saturation. This is also advantageous because this is the “unloading” portion
of the curve where hemoglobin loses its oxygen to the tissues.
Many factors determine the hemoglobin
saturation. If, for example, the
blood becomes more acidic, the dissociation curve shifts to the right. This
indicates that more oxygen is being unloaded from the hemoglobin at the tissue
level. This rightward shift of the curve(figure a), attributable to a decline in pH, is
reffered to as the Bohr effect. The pH in the lungs is generally high, so
hemoglobin passing through the lungs has a strong affinity for oxygen,
encouraging high saturation. At the tissue level, especially during exercise,
the pH is lower, causing oxygen to dissociate from hemoglobin, thereby
supplying oxygen to the tissues. With exercise, the ability to unload oxygen to
the muscles increases as the muscle pH decreases.
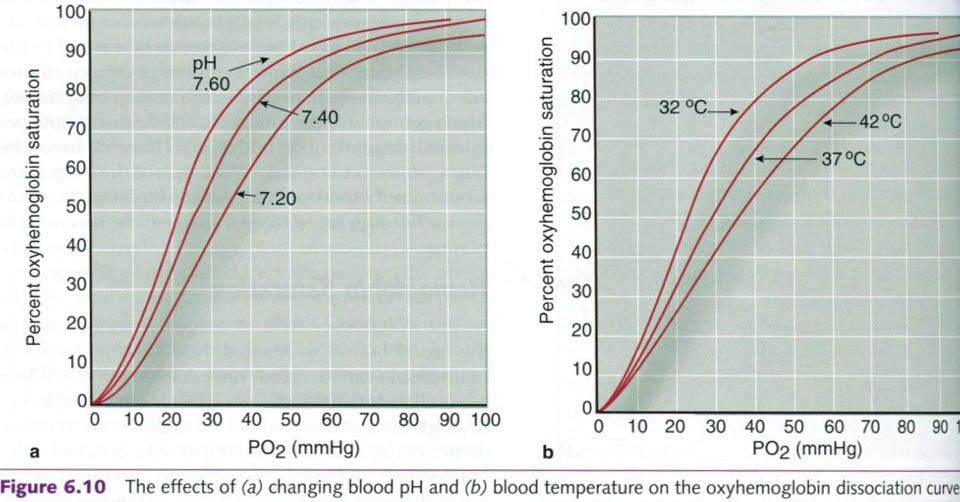
Blood temperature also affects oxygen
dissociation. As shown in the figure b, increased blood temperature shifts the
dissociation curve to the right, indicating that oxygen is unloaded from
hemoglobin more readily at higher temperatures. Because of this, the hemoglobin
unloads more oxygen when blood circulates through the metabolically heated
active muscles.
Blood oxygen-carrying capacity
The oxygen-carrying capacity of blood is the
maximal amount of oxygen the blood can transport. It depends primarily on the
blood hemoglobin content. Each 100ml of blood contains an average of 14 to 18g
or hemoglobin in men and 12 to 16g in women. Each gram of hemoglobin can
combine with about 1.34ml of oxygen, so the oxygen-carrying capacity of blood
is approximately 16 to 24ml per 100ml of blood when blood is fully saturated
with oxygen. At rest, as the blood passes through the lungs, it is in contact
with the alveolar air for approximately 0.75s. This is sufficient time for
hemoglobin to become 98% to 99% saturated. At high intensities of exercise, the
contact time is greatly reduced, which can reduce the binding of hemoglobin to
oxygen and slightly decrease the saturation, although the unique S shape of the
curve guards against large drops.
People with low hemoglobin concentrations, such
as those with anemia, have reduced oxygen-carrying capacities. Depending on the
severity of the condition, these people might feel few effects of anemia while
they are at rest because their cardiovascular system can compensate for reduced
blood oxygen content by increasing cardiac output. However, during activities
in which oxygen delivery can become a limitation, such as highly intense
aerobic effort, reduced blood oxygen content limits performance.
Carbon dioxide transport
Carbon dioxide also relies on the blood for
transportation. Once carbon dioxide is released from the cells, it is carried
in the blood primarily in three forms:
- As bicarbonate ions
resulting from the dissociation of carbonic acid
- Dissolved in plasma
- Bound to hemoglobin(called
carbaminohemoglobin).
Bicarbonate ion
The majority of carbon dioxide is carried in
the form of bicarbonate ion. Bicarbonate accounts for the transport of 60% to
70% of the carbon dioxide in the blood. Carbon dioxide and water molecules
combine to form carbonic acid(H2CO3). This reaction is
catalyzed by the enzyme carbonic anhydrase, which is found in red blood cells.
Carbonic acid is unstable and quickly dissociates, freeing a hydrogen ion(H+)
and forming a bicarbonate ion(HCO3-):
CO2 + H2O --> H2CO3 --> H+ + HCO3-
The H+ subsequently binds to
hemoglobin, and this binding triggers the Bohr effect, mentioned previously,
which shifts the oxygen-hemoglobin dissociation curve to the right. The bicarbonate
ion diffuses out of the red blood cell and into the plasma. In order to prevent electrical imbalance from
the shift of the negatively charged bicarbonate ion into the plasma, a chloride
ion diffuses from the plasma into the red blood cell. This is called the
chloride shift.
Additionally, the formation of hydrogen ions
through this reaction enhances oxygen unloading at the level of the tissue.
Through this mechanism, hemoglobin acts as a buffer, binding and neutralizing
the H+ and thus preventing any significant acidification of the blood.
When the blood enters the lungs, where the PCO2
is lower, the H+ and bicarbonate ions rejoin to form carbonic acid, which then
dissociates into carbon dioxide and water:
H+ + HCO3- --> H2CO3 --> CO2 + H2O
The carbon dioxide that is thus re-formed can
enter the alveoli and be exhaled.
Dissolved carbon dioxide
Part of the carbon dioxide released from the
tissues is dissolved in plasma; but only a small amount, typically just 7% to
10%, is transported this way. This dissolved carbon dioxide comes out of
solution where the PCO2 is low, as in the lungs. There it diffuses
from the pulmonary capillaries into the alveoli to be exhaled.
Carbaminohemoglobin
Carbon
dioxide transport also can occur when the gas binds with hemoglobin, forming
carbaminohemoglobin. The
compound is so named because carbon dioxide binds with amino acids in the
globin part of the hemoglobin molecule, rather than with the heme group as
oxygen does. Because carbon dioxide binding occurs on a different part of the
hemoglobin molecule than does oxygen binding, the two processes do not compete.
However, carbon dioxide binding varies with the oxygenation of the
hemoglobin(deoxyhemoglobin binds carbon dioxide more easily than oxyhemoglobin)
and the partial pressure of CO2. Carbon dioxide is released from
hemoglobin when PCO2 is low as it is in the lungs. Thus, carbon
dioxide is readily released from the hemoglobin in the lungs, allowing it to
enter the alveoli to be exhaled.












0 коментара:
Постави коментар