



Regulation of glucose metabolism during exercise
The heightened energy demands of exercise
require that more glucose be made available to the muscles. Recall that glucose
is stored in the body as glycogen,
primarily in the muscles and the liver. Glucose must be freed from its storage
form of glycogen, so glycogenolysis must increase. Glucose freed from the liver
enters the blood to circulate throughout the body, allowing it access to active
tissues. Plasma glucose concentration also can be increased through gluconeogenesis.
Regulation of plasma glucose concentration
Four hormones work to increase the amount of
circulating plasma glucose:
- Glucagon
- Epinephrine
- Norepinephrine
- Cortisol
The plasma glucose concentration during
exercise depends on a balance between glucose uptake by exercising muscles and
its release by the liver. At rest, glucose release from the liver is
facilitated by glucagon, which promotes both liver glycogen breakdown and
glucose formation from amino acids. During exercise, glucagon secretion
increases. Muscular activity also increases the rate of catecholamine release
from the adrenal medulla, and these
hormones(epinephrine and norepinephrine) work with glucagons to further
increase glycogenolysis. Cortisol concentrations also increase during exercise.
Cortisol increases protein
catabolism, freeing amino acids to be used within the liver for gluconeogenesis. Thus, all four of these
hormones can increase plasma glucose by enhancing the processes of glycogenolysis(breakdown
of glycogen) and gluconeogenesis(making glucose from other substrates). In
addition to the effects of the four major glucose-controlling hormones, growth hormone increases mobilization of
FFAs and decreases cellular uptake of glucose, so less glucose is used by the
cells(more remains in circulation); and the thyroid hormones promote glucose
catabolism and fat metabolism.
The amount of glucose released by the liver
depends on exercise intensity and the duration. As intensity increases, so does
the rate of catecholamine release. This can cause the liver to release more
glucose than is being taken up by the active muscles. Consequently, during or
shortly after an explosive, short-term sprint bout, blood glucose
concentrations may be 40% to 50% above the resting level, illustrating that the
glucose release by the liver is greater than the uptake by the muscles.
The greater the exercise intensity, the greater
the catecholamine release, and thus the glycogenolysis rate is significantly
increased. This process occurs not only in the liver but also in the muscle.
The glucose released from the liver enters the blood to become available to the
muscle. But the muscle has a more readily available source of glucose: its own
glycogen. The muscle uses its own glycogen stores before using the plasma
glucose during explosive, short-term exercise. Glucose released from the liver
is not used as readily, so it remains in the circulation, elevating the plasma
glucose. Following exercise, plasma glucose concentrations decrease as the
glucose enters the muscle to replenish the depleted muscle glycogen stores.
During exercise bouts that last for several
hours, however, the rate of liver glucose release more closely matches the
muscles’ needs, keeping plasma glucose at or only slightly above the resting
concentrations. As muscle uptake of glucose increases, the liver’s rate of
glucose release also increases. In most cases, plasma glucose does not begin to
decline until late in the activity as liver glycogen stores become depleted, at
which time glucagons concentrations increase significantly. Glucagon and cortisol together enhance gluconeogenesis,
providing more fuel.
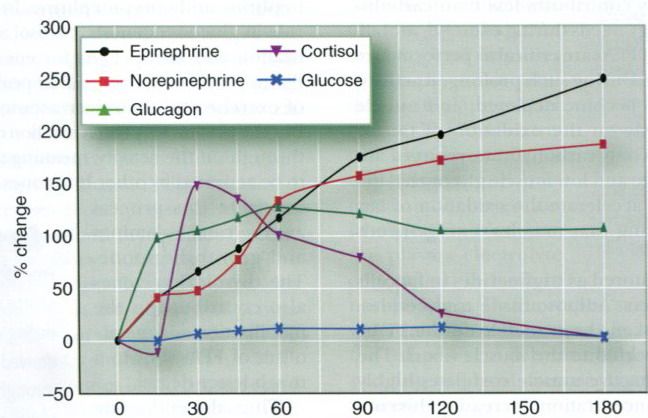
Figure below illustrates the changes in plasma
concentrations of epinephrine, norepinephrine, glucagon, cortisol, and glucose
during 3h of cycling. Although the hormonal regulation of glucose remains
intact throughout such longterm activities, the liver’s glycogen supply may
become critically low. As a result, the liver’s rate of glucose release may be
unable to keep pace with the muscles’ rate of glucose uptake. Under this
condition, plasma glucose may decline despite strong hormonal stimulation.
Glucose ingestion during the activity can play a major role in maintaining
plasma glucose concentrations.
Glucose uptake by muscle
Merely releasing sufficient amounts of glucose
into the blood does not ensure that the muscle cells will have enough glucose
to meet their energy demands. Not only must the glucose be released and
delivered to these cells; it also must be taken up by the cells. Transport of
glucose through the cell membranes and into muscle glucose through the cell
membranes and into muscle cells in controlled by insulin. Once glucose is delivered to the muscle, insulin
facilitates its transport into the fibers.
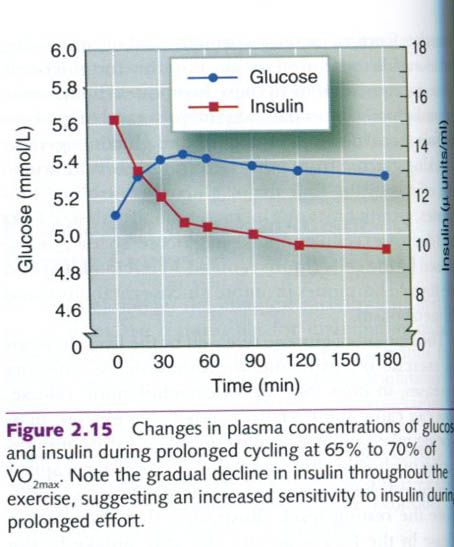
Surprisingly, as seen in figure below, plasma
insulin concentration tends to decrease during prolonged submaximal exercise,
despite a slight increase in plasma glucose concentration and glucose uptake by
muscle. This apparent contradiction between the plasma insulin concentrations
and the muscles’ need for glucose serves as a reminder that a hormone’s
activity is determined not only by its concentration in the blood but also by a
cell’s sensitivity to that hormone. Exercise may enhance insulin’s binding to
receptors on the muscle fiber, thereby reducing the need for high
concentrations of plasma insulin to transport glucose across the muscle cell
membrane into the cell. This is important, because during exercise four
hormones are trying to release glucose from its storage sites and create new
glucose. High insulin concentrations would oppose their action, preventing this
needed increase in plasma glucose supply.
Regulation of fat metabolism during exercise
Although fat generally contributes less than
carbohydrate to muscles’ energy needs during exercise, mobilization and
oxidation of FFAs are critical to performance in endurance exercise. During
such prolonged activity, carbohydrate reserves become depleted, and muscle must
rely more heavily on the oxidation of fat for energy production. When
carbohydrate reserves are low(low plasma glucose and low muscle glycogen), the
endocrine system can accelerate the oxidation of fats(lypolysis), thus ensuring
that muscles’ energy needs can be met.
Free fatty acids are stored as triglycerides in
fat cells and inside muscle fibers. Adipose tissue triglycerides, however, must
be broken down to release the FFAs, which are then transported to the muscle
fibers. The rate of FFA uptake by active muscle correlates highly with the
plasma FFA concentration. Increasing this concentration would increase cellular
uptake of the FFA. The rate of triglyceride breakdown may determine, in part,
the rate at which muscles use fat as a fuel source during exercise.
The rate of lypolysis is controlled by at least
five hormones:
- (decreased) Insulin
- Epinephrine
- Norepinephrine
- Cortisol
- Growth hormone
The major factor responsible for adipose tissue
lypolysis during exercise is a fall in circulating insulin. Lipolysis is also
enhanced through the elevation of epinephrine and norepinephrine. In addition
to having a role in gluconeogenesis,
cortisol accelerates the mobilization and use of FFAs for energy during
exercise. Plasma cortisol concentration peaks after 30 to 45 min of exercise
and then decreases to near-normal levels. But the plasma FFA concentration
continues to increase throughout the activity, meaning that lipase continues to
be activated by other hormones. The hormones that continue this process are the
catecholamines and growth hormone. The thyroid hormones also contribute to the
mobilization and metabolism of FFAs, but to a much lesser degree.
Thus, the endocrine system plays a critical
role in regulating ATP production during exercise as well as controlling the
balance between carbohydrate and fat metabolism.
“Physiology of sport and exercise”, fourth
edition; Jack H. Wilmore, David L. Costill, W. Larry Kenney








2 коментара:
No one can have their perfect diet which includes all minerals and vitamins. Sometimes we need to have some supplements to balance the nutrients in our body. Supplementation may help you live longer. To know more about supplements please check this out--> Natural Sleep Aid
Growth hormone plays an important role in maintaining a lean physique. When the body stops producing HGH, the first signs of aging become obvious. For many men, these first signs include the loss of muscle mass and the accumulation of belly fat.
An increase in growth hormone can slow down the hands of time. Apart from affecting muscle mass and the body fat percentage, HGH does a number of other important things. It makes the skin healthy, improves bone density and even has a positive impact on sleep quality.
compelling vigrxplus website
Постави коментар