



Adequate oxygen supply to exercising muscles is
essential to physical performance and it depends on an adequate supply of
oxygen being brought into the body, moved
from the lungs to the blood,
transported to the muscles, and adequately taken up into the muscles. A
limitation in any of these steps can impair performance.
The transport of oxygen to working muscle
begins with pulmonary ventilation,
the active transport of gas molecules into the alveoli of the lungs(breathing).
Ventilation increases within seconds of exposure to high altitude, both at rest
and during exercise, because chemoreceptors in the aortic arch and carotid
arteries are stimulated by the low PO2 and signals are sent to the
brain to increase breathing. The increased ventilation is primarily associated
with an increased tidal volume,
although respiratory rate also increases. Over the next several hours and days,
ventilation remains elevated to a level proportional to the altitude.
Increased ventilation acts much the same as
hyperventilation at sea level. The amount of carbon dioxide in the alveoli is
reduced. Carbon dioxide follows the presture gradient, so more diffuses out of
the blood, where its pressure is
relatively high, and into the lungs to be exhaled. This “blowing off” of CO2
causes blood PCO2 to fall and blood pH to increase, a condition
known as respiratory alkalosis. This
alkalosis has two effects. First, it causes the oxyhemoglobin saturation curve
to shift to the left. Second, it helps limit the rise in ventilation but is
overridden by the hypoxic(low PO2) drive. At a given submaximal
exercise intensity, ventilation is higher at altitude than at sea level, but maximal
exercise ventilation is similar.
In an effort to offset respiratory alkalosis,
the kidneys excrete more bicarbonate ion, the ions that buffer the carbonic
acid formed from carbon dioxide. Thus, a reduction in bicarbonate ion
concentration reduces the blood’s buffering capacity. More acid remains in the blood, and the alkalosis is offset.
Under resting conditions, pulmonary diffusion(diffusion of O2 from the alveoli to
the arterial blood) does not limit the exchange of gases between the alveoli
and the blood. If gas exchange were
limited or impaired, less oxygen would enter the blood, so the arterial PO2 would be much lower than the
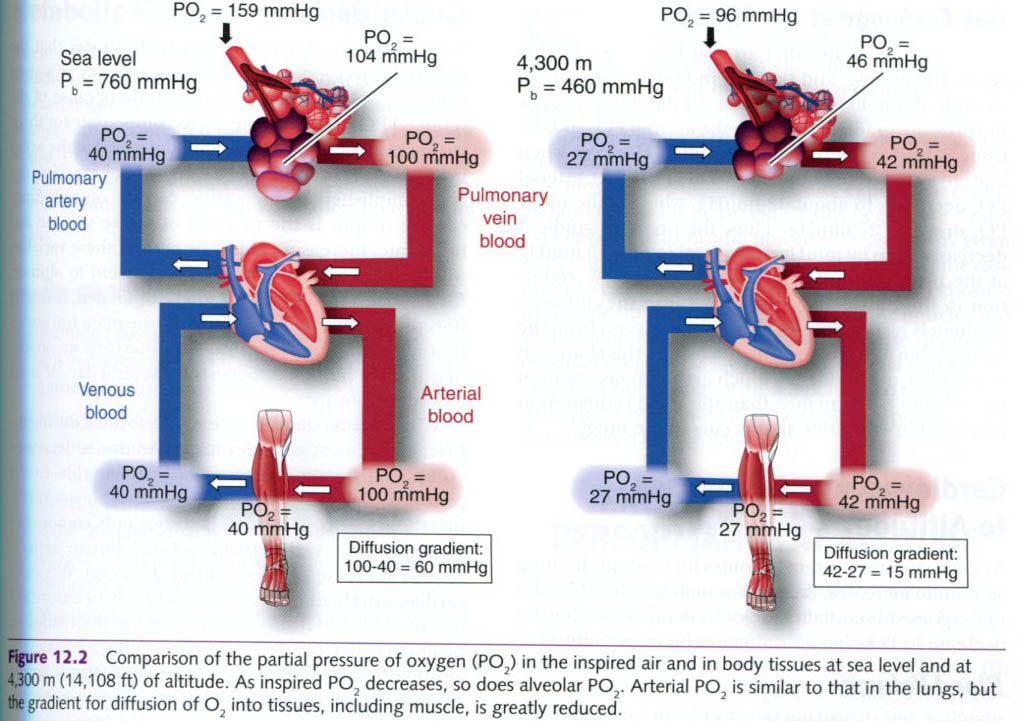
alveolar PO2. Instead, these two values are almost equal(see picture
below). Therefore, the low arterial blood PO2, or hypoxemia, is a
direct reflection of the low alveolar PO2, and not an oxygen
diffusion limitation from the lungs to the blood.
Oxygen
transport
As shown in the figure above, the PO2
at sea level is 159mmHg; however, it decreases to about 104 in the alveoli primarily because of
the addition of water vapor molecules(PH2O = 47mmHg at 37°C). When
the alveolar PO2 drops at altitude, less hemoglobin in the blood
perfusing the lungs becomes saturated with O2. As depicted in the
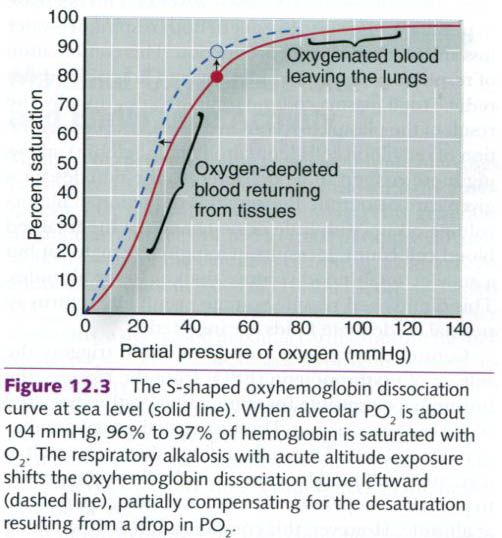
figure below, the oxygen-binding curve for hemoglobin has a distinct S shape. At
sea level, when alveolar PO2 is about 104mmHg, 96% to 97% of
hemoglobin has O2 bound to it. When PO2 in the lungs is
decreased to 46mmHg at 4,300m(14,108ft), only about 80% of hemoglobin sites are
saturated with O2. If the oxygen-loading portion of the curve were
not relatively flat, far less O2 would be taken up by the blood as
it passes through the lungs. Therefore, while arterial blood is still
desaturated at altitude, the features of the oxyhemoglobin dissociation curve
serve to minimize this problem.
A second adaptation occurs very early in
altitude exposure that also aids in preventing the fall in arterial oxygen
content. As mentioned earlier, a respiratory alkalosis accompanies the
increased ventilation caused by acute altitude exposure. This increase in blood
pH actually shifts the oxyhemoglobin dissociation curve leftward, as shown in
the figure below. The result is that, rather than 80% binding of oxygen to
hemoglobin, 89% of Hb is saturated with O2.
Gas exchange in the muscles
First figure shows that arterial PO2
at sea level is about 100mmHg, and the PO2 in body tissues is
consistently about 40mmHg at rest; so the difference, or the pressure gradient,
between the arterial PO2 and the tissue PO2 at sea level
is about 60mmHg. However, when one moves to an elevation of 4,300m(14,108ft),
arterial PO2 decreases to about 42mmHg, whereas the tissue PO2
drops to 27mmHg. Thus, the pressure gradient decreases from 60mmHg at sea level
to only 15mmHg at the higher altitude. This represents a 75% reduction in the
diffusion gradient! Because the diffusion gradient is responsible for driving
the oxygen from the hemoglobin in the blood into the tissues, this change in
arterial PO2 at altitude is a much greater consideration for
exercise performance than the small reduction in hemoglobin saturation that
occurs in the lungs.
Cardiovascular responses to altitude
As the respiratory system becomes increasingly
limiting as altitude increases, the cardiovascular system likewise undergoes
substantial changes to compensate for the decrease in PO2 that
accompanies increased altitude.
Blood
volume
Within the first few hours of arrival at
altitude, a person’s plasma volume begins to progressively decrease, and it
plateaus by the end of the first few weeks. This decrease in plasma volume is
the result of both respiratory water loss and increased urine production. This
combination of respiratory water loss and the excretion of fluid can reduce
total plasma volume by up to 25%. Initially, the result of the plasma loss is
an increase in the concentration of red blood cells(containing hemoglobin),
allowing more oxygen to be delivered to the muscles for a given cardiac output.
Because this reduction in plasma volume occurs with little or no change in the
total red blood cell count, it results in a higher hematocrit but a smaller
total blood volume than at lower altitudes. This diminished plasma volume
eventually returns to normal if adequate fluids are ingested.
Continued exposure to high altitude triggers
the release of erythropoetin(EPO) from the kidneys, the hormone responsible
for stimulating erythrocyte(red blood cell) production. This increases the
total number of red blood cells. These adaptations ultimately result in a
greater total blood volume, which
allows the person to partially compensate for the lower PO2
experienced at altitude. However, this compensation is slow, taking weeks to
months to fully restore red cell mass.
Cardiac
output
The preceding discussion clearly illustrates
that the amount of oxygen delivered to the muscles by a given volume of blood is limited at altitude because of
the reduced PO2. A logical means to compensate for this is to
increase the volume of blood delivered to the active muscles. At rest and
during submaximal exercise, this is accomplished by increasing cardiac output. Upon ascent to altitude,
the sympathetic nervous system is stimulated, releasing norepinephrine and epinephrine,
the main hormones that alter cardiac function. The increase in norepinephrine in particular persists for several
days of acute altitude exposure.
When submaximal exercise is performed during
the first few hours at altitude, stroke volume is decreased compared to sea-level values(attributable to the
reduced plasma volume). Fortunately, heart rate is increased disproportionately to not only compensate for the
decrease in stroke volume but to
actually slightly increase cardiac output.
However, this extra cardiac workload is not an efficient way to ensure
sufficient oxygen delivery to the body’s active tissues for prolonged periods.
Consequently, after a few days at altitude, the muscles begin extracting more
oxygen from the blood(increasing the
arterial-venous oxygen difference), which reduces the demand for increased
cardiac output, since VO2 = Q x (a-ṽ)O2 difference. The
increase in heart rate and cardiac output peaks after about 6 to 10 days at
high altitude, after which cardiac output and heart rate during a given
exercise bout start to decrease.
At maximal or exhaustive work levels at higher
altitudes, both maximal stroke volume
and maximal heart rate are decreased.
The decrease in stroke volume is
directly related to the decrease in plasma volume. Maximal heart rate may be somewhat lower at high altitude as a consequence
of a decrease in the response to sympathetic nervous system activity, possibly
attributable to a reduction in ß-receptors(receptors in the heart that respond to sympathetic nerve
activation, thus increasing the heart rate). The effect of these changes in maximal heart rate and stroke volume
is obvious: maximal cardiac output decreases. With a decreased diffusion
gradient to push oxygen from the blood
into the muscles, coupled with this reduction in maximal cardiac output, it is apparent why both VO2max and
submaximal aerobic performance are hindered at altitude. In summary, hypobaric
conditions significantly limit oxygen delivery to the muscles, reducing the
capacity to perform high-intensity or prolonged aerobic activities.
Metabolic responses to altitude
Ascent to altitude increases the basal
metabolic rate, possibly due to increases in either or both thyroxin and the catecholamines. This increased metabolism must be balanced by an
increased food intake to prevent body weight from decreasing, a common
occurrence during the first few days at altitude because appetite declines as
well. In individuals who maintain their body weight at altitude, there is an
increased reliance on carbohydrate
for fuel, both at rest and during submaximal exercise. Because glucose yields
more energy than fats or proteins per liter of oxygen, this
adaptation would be beneficial.
Given the hypoxic conditions at altitude, and
because any fixed amount of work at altitude represents a higher percentage of
VO2max, we would expect anaerobic metabolism to be increased. If
this occurs, we would expect lactic acid production to increase at any given
work rate above lactate threshold.
This is in fact the case upon arrival at altitude. However, with longer
exposure to altitude the lactate concentration in the muscles and venous blood
at a given intensity of exercise(including maximal exertion) is lower, despite
the fact that muscle VO2 does not change with adaptation to
altitude. To date, there is no universally accepted explanation for this
so-called lactate paradox.











0 коментара:
Постави коментар