



Blood flow
Acute changes in cardiac output and blood
pressure during exercise allow for increased total blood flow to the body.
These responses facilitate getting blood to areas where it is needed, primarily
the exercising muscles. Additionally, sympathetic control of the cardiovascular
system can redistribute blood so that areas with the greatest metabolic need
receive more blood than areas with low demands.
Redistribution
of blood during exercise
Blood flow patterns change markedly in the
transition from rest to exercise. Through the action of the sympathetic nervous
system, blood is redirected away from areas where elevated flow is not
essential to those areas that are active during exercise. Only 15% to 20% of
the resting cardiac output goes to muscle, but during high-intensity exercise,
the muscles may receive 80% to 85% of the cardiac output. This shift in blood
flow to the muscles is accomplished primarily by reducing blood flow to the
kidneys and the primarily by reducing blood flow to the kidneys and the
so-called splanchnic circulation that includes the liver, stomach, and
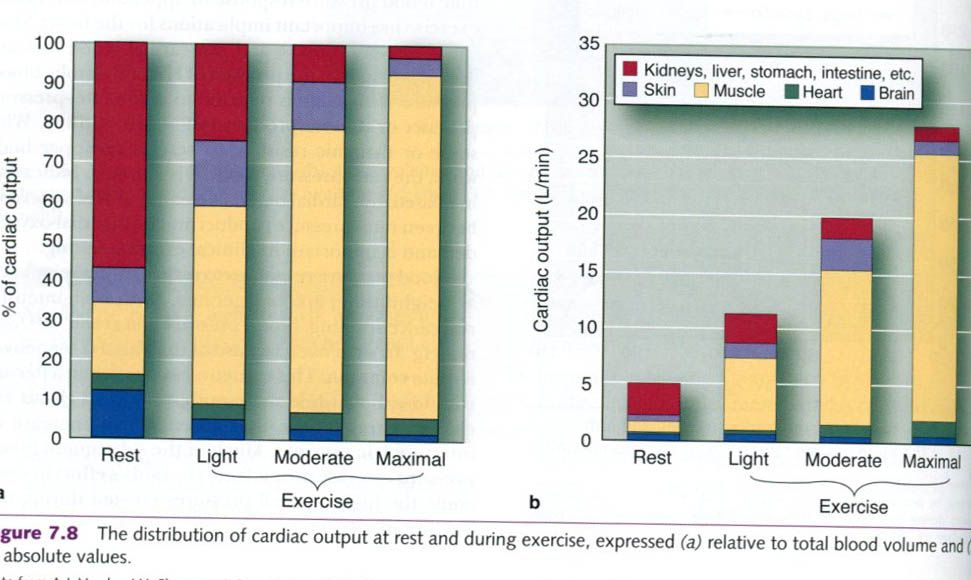
intestines. Figure below illustrates a typical distribution of cardiac output
throughout the body at rest and during heavy exercise. Because cardiac output
increases greatly with increasing intensity of exercise, the values are shown
both as relative percentages of the total blood available and as absolute
flows.
Although several physiological mechanisms are
responsible for the redistribution of blood flow through the body during
exercise, they work together. To illustrate this, consider what happens to
blood flow during exercise, focusing on the primary driver of the response,
namely the blood flow requirements of the skeletal muscles.
As exercise begins, the active skeletal muscles
rapidly sense the need for increased oxygen delivery. This need is met through
sympathetic stimulation of vessels in those areas to which blood flow is to be
reduced( e.g. the splanchnic and renal circulations). The resulting
vasoconstriction in those areas allows for more of the increasing cardiac
output to be redistributed to the exercising skeletal muscles. In the skeletal
muscles, sympathetic stimulation to the constrictor fibers in the arteriolar
walls also increases; however local vasodilating substances are released from
the exercising muscle and overcome sympathetic vasoconstriction, producing an
overall vasodilation in the muscle.
Many local vasodilating substances are released
in exercising skeletal muscle. As the metabolic rate of the muscle tissue
increases during exercise, metabolic waste products begin to accumulate.
Increased metabolism causes an increase in acidity(increased hydrogen ions and
lower pH), carbon dioxide, and temperature in the muscle tissue. These are some
of the local changes that trigger vasodilation of, and increasing blood flow
through, the arterioles feeding local capillaries. Local vasodilation is also
triggered by the low partial pressure of oxygen in the tissue or a reduction in
oxygen bound to hemoglobin(increased oxygen demand), the act of muscle
contraction, and possibly other vasoactive substances(including adenosine)
released as a result of skeletal muscle contraction.
When exercise is performed in a hot
environment, there is an increase in blood flow to the skin to help dissipate
the body heat. The sympathetic control of skin blood flow is unique in that
there are both typical sympathetic vasoconstrictor fibers(similar to skeletal
muscle) and sympathetic active vasodilator fibers interacting. During dynamic
exercise, skin blood flow is initially and transiently decreased by an increase
in sympathetic vasoconstrictor activity. As body core temperature rises, there
is a reduction in this sympathetic vasoconstriction, causing a passive
vasodilation. Finally, at a specific body core temperature threshold, skin
blood flow begins to dramatically increase through the action of the
sympathetic active vasodilator system. The increase in skin blood flow during
exercise promotes heat loss, because metabolic heat from deep in the body can
be released when blood moves close to the skin. This allows maintenance of the
body temperature, although body temperature does increase with exercise.
Cardiovascular
drift
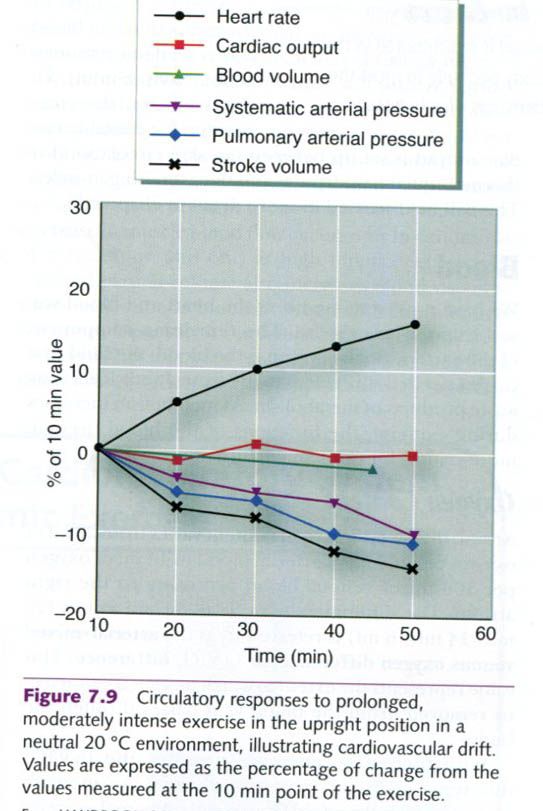
With prolonged aerobic exercise or aerobic
exercise in a hot environment, at a constant exercise intensity, SV gradually
decreases and HR increases. Cardiac output is well maintained, but arterial
blood pressure also declines. These alterations, illustrated in the figure
below, have been referred to collectively as cardiovascular drift, and they are generally associated with
increasing body temperature. Cardiovascular drift is associated with a
progressive increase in the fraction of cardiac output directed to the
vasodilated skiin to facilitate heat loss and attenuate the increase in body
core temperature. With more blood in the skin for the purpose of cooling the
body, less blood is available to return to the heart, thus decreasing preload.
There is also a small decrease in blood volume resulting from sweating and from
a generalized shift of plasma across the capillary membrane into the
surrounding tissues. These factors combine to decrease ventricular filling
pressure, which decreases venous return to the heart and reduces the
end-diastolic volume. With the reduction in end-diastolic volume, SV is
reduced(SV = EDV – ESV). In order to maintain cardiac output(Q = HR x SV). HR
increases to compensate for the decrease in SV.
Competition
for blood supply
When the demands of exercise are added to blood
flow demands for all other systems of the body, competition for a limited
available cardiac output can occur. This competition for available blood flow
can develop among several vascular beds, depending on the specific conditions.
For example, there is a competition for blood flow between active skeletal
muscle and the gastrointestinal system following a meal. McKirnan and coworkers
studied the effects of feeding versus fasting on the distribution of blood flow
during exercise in miniature pigs. The pigs were divided into two groups. One
group fasted for 14 to 17h before exercise. The other group ate their morning
ration in two feedings: half the ration was fed 90 to 120 minutes before
exercise and the other half 30 to 45 minutes before exercise. Both groups of
pigs then ran at approximately 65% of their VO2max.
Blood flow to the hindilimb muscles during
exercise was 18% lower, and gastrointestinal blood flow was 23% higher, in the
fed group than in the fasted group. Similar results in humans suggest that the
redistribution of gastrointestinal blood flow to the working muscles is
attenuated after a meal. As a practical application, these findings suggest
that athletes should be cautious in timing their meals before competition to
maximize blood flow to the active muscles during exercise.
Another example of the competition for blood
flow is seen in exercise in a hot environment. In this scenario, the
competition for available cardiac output is set up between the skin circulation
for thermoregulatory purposes and the exercising muscles.
The remaining component is blood: the fluid
that carries needed substances to the tissues and clears away waste products of
metabolism. As metabolism increases during exercise, the functions of the blood
become more critical for optimal performance.
Oxygen
content
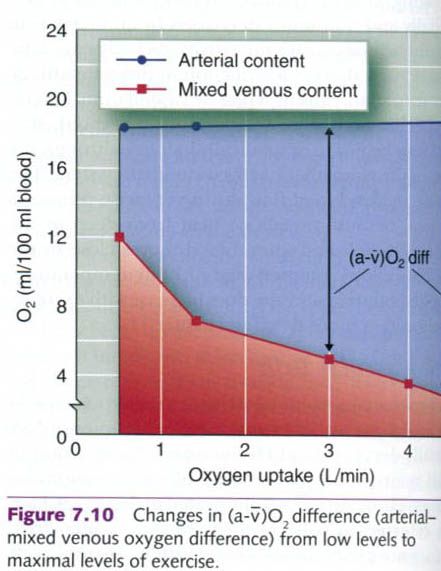
At rest, the blood’s oxygen content varies from
20ml of oxygen per 100ml of arterial blood to 14ml of oxygen per 100ml of
venous blood returning to the right atrium. The difference between these two
values(20ml – 14ml = 6ml) is referred to as the arterial-mixed venous oxygen
difference, or (a-ṽ)O2 difference. This value represents the extent
to which oxygen is extracted, or removed, from the blood as it passes through
the body.
With increasing exercise intensity, the (a-ṽ)O2
difference increases progressively and can increase approximately threefold
from rest to maximal exercise intensities(figure below). This increased
difference really reflects a decreasing venous oxygen content, because arterial
oxygen content changes little from rest up to maximal exertion. With exercise,
more oxygen is required by the active muscles: therefore more oxygen is
extracted from the blood. The venous oxygen content decreases, approaching zero
in the active muscles. However, mixed venous blood in the right atrium of the
heart rarely decreases below 4ml of oxygen per 100ml of blood because the blood
returning from the active tissues is mixed with blood from inactive tissues as
it returns to the heart. Oxygen use in the inactive tissues is far lower than
in the active muscles.
Plasma
volume
With the onset of exercise, there is an almost
immediate loss of plasma from the blood to the interstitial fluid space. The
movement of fluid out of the capillaries is dictated by the pressures inside
the capillaries, which include the hydrostatic or blood pressure and the
osmotic pressure exerted by the proteins in the blood, mostly albumin. The pressures that influence fluid movement
outside the capillaries are the pressure provided by the surrounding tissue as
well as the osmotic pressures from proteins in the interstitial fluid(figure
below). As blood pressure increases with exercise, the hydrostatic pressure
within the capillaries also increases. Thus, the increase in blood pressure
forces water from the vascular compartment to the interstitial compartment.
Also, as metabolic waste products build up in the active muscle, intramuscular
osmotic pressure increases, which draws fluid out of the capillaries to the
muscle.

Approximately a 10% to 15% reduction in plasma
volume can occur with prolonged exercise. Similarly, 15% to 20% decreases in
plasma volume have been observed in 1 min bouts of exhaustive exercise. With
resistance training, the plasma volume loss is proportional to the intensity of
the effort, with losses of from 10% to 15%.
If exercise intensity or environmental
conditions cause sweating, additional plasma volume losses may occur. Although
the major source of fluid for sweating is the interstitial fluid, this fluid
space will be diminished as sweating continues. This increases the osmotic
pressure in the interstitial space(since proteins do not move with the fluid),
causing even more plasma to move out of the vascular compartment into the
interstitial space. Intracellular fluid volume is impossible to measure
directly and accurately, but research suggests that fluid is also lost from the
intracellular compartment and even from the red blood cells, which may shrink
the size.
A reduction of plasma volume will impair
performance under many circumstances. For long-duration activities in which
dehydration occurs and heat loss is a problem, the total flow of blood to
active tissues must be reduced to allow increasingly more blood to be diverted
to the skin in an attempt to lose body heat. Note that a decrease in muscle
blood flow occurs only in conditions of dehydration and only at high
intensities. Severely reduced plasma volume also increases blood viscosity,
which can impede blood flow and thus limit oxygen transport, especially if the
hematocrit exceeds 60%.
In activities that last a few minutes or less,
body fluid shifts are of little practical importance. As exercise duration
increases, however, body fluid changes and temperature regulation become
important for performance. For the football player, the Tour de France cyclist,
or the marathon runner, these processes are crucial, not only for competition
but also for survival. Deaths have occurred from dehydration and hyperthermia
during or as a result of various sport activities.
Hemoconcentration
When plasma volume is reduced, hemoconcentration occurs: the fluid
portion of the blood is reduced, and the cellular and protein portions
represent a larger fraction of the total blood volume. That is, they become
more concentrated in the blood. This hemoconcentration increases red blood cell
concentration substantially – by up to 20% or 25%. Hematocrit can increase from
40% to 50%. However, the total number and volume of red blood cells do not
change substantially.
The net effect, even without an increase in the
total number of red blood cells, is to increase the number of red blood cells
per unit of blood; that is, the cells are more concentrated. As the red blood
cell concentration increases, so does the blood’s per unit hemoglobin content. This
substantially increases the blood’s oxygen-carrying capacity, which is
advantageous during exercise and provides a distinct advantage at altitude at
rest and during submaximal exercise.
Integration of the exercise response
As is evident from all of the changes in
cardiovascular function that take place at the onset of exercise, the
cardiovascular system is extremely complex but responds exquisitely to deliver
oxygen to meet the demands of exercising muscle. Figure below is a simplified
flow diagram that illustrates how the body integrates all these cardiovascular
responses to provide for its needs during exercise. Key areas and responses are
labeled and summarized to help illustrate how these complex control mechanisms
are coordinated. It is important to note that although the body attempts to
meet the blood flow needs of the muscle, it can do so only if blood pressure is
not compromised. Maintenance of arterial blood pressure appears to be the
highest priority of the cardiovascular system, irrespective of exercise, the
environment, and other competing needs.
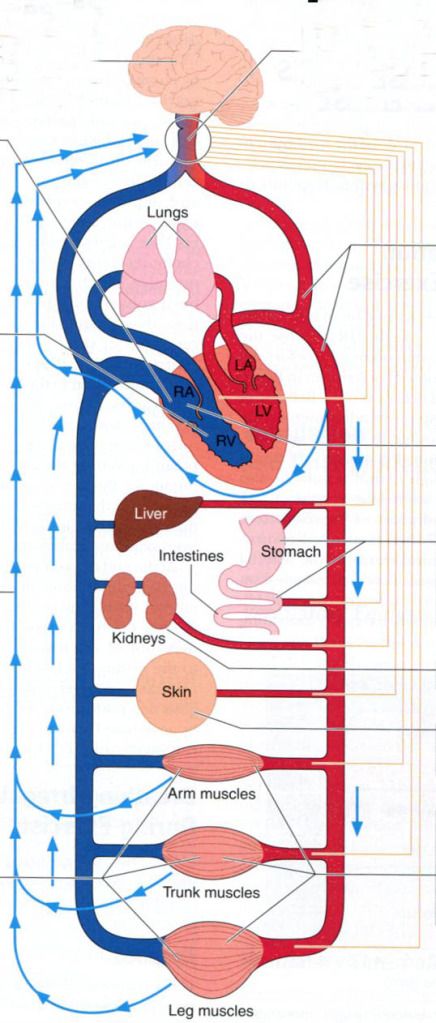
1) Cardiovascular
function is regulated by the medulla in the brain.
2) Medulla
regulates heart rate via the pacemaker in the right atrium.
3) Right
atrium is stimulated by sympathetic and parasympathetic nerves to raise and
lower heart rate.
4) The
motor cortex of the brain stimulates the medulla in proportion to the amount of
muscle recruited.
5) Sensory
nerves in the muscle send impulses to the medulla regarding the metabolic
status of the muscle.
6) High-pressure
baroreceptors in the arteries send impulses to the medulla regarding arterial
blood pressure.
7) With
increases in exercise intensity, sympathetic nerves cause reduction of blood
flow to the stomach, liver, and intestines…
8) …
and the kidneys…
9) …
and the skin(unless there is a need for heat dissipation).
10) Release
of metabolites from exercising skeletal muscles causes vasodilation of the
arterioles leading to the active fibers.
11) However,
muscle can accept too much blood flow; thus sympathetic nerves to exercising
muscles must increase resistance somewhat to maintain blood pressure.
12) Low-pressure
baroreceptors in the right side of the heart and pulmonary circulation detect
the extent to which the heart is filling with blood. With increased filling,
the low-pressure baroreceptors send impulses to the brain to lower resistance
to exercising skeletal muscles(11); thus, arterial pressure is maintained.




0 коментара:
Постави коментар