



31. 5. 2012.
Neuromuscular junctions and neurotransmitters
Neuromuscular junction
Whereas neurons communicate with other neurons
at synapses, an alpha-motor neuron communicates with muscle fibers at a site
known as a neuromuscular junction.
The function of the neuromuscular junction is essentially the same as that of a
synapse. In fact, the proximal part of the neuromuscular junction is the same: It
starts with the axon terminals of the motor neuron, which release
neurotransmitters into the space between the motor nerve and the muscle fiber
in response to an action potential. However, in the neuromuscular junction,
the axon terminals protrude into motor end plates, which are troughlike
segments on the plasmalemma. Picture below shows.
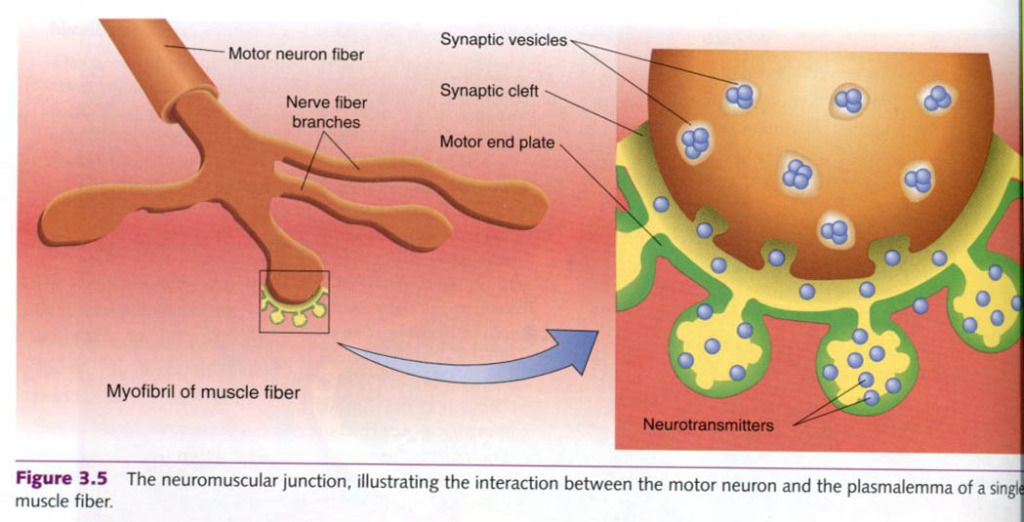
The motor end plate is invaginated(folded to
form cavities). The cavity thus formed is called the synaptic gutter. As with
synapses, the space between the neuron and the muscle fiber is the synaptic cleft.
Neurotransmitters
released from the alpha-motor neuron axon terminals diffuse across the synaptic
cleft and bind to receptors on the muscle fiber’s plasmalemma. This binding
typically causes depolarization by opening sodium ion channels, allowing more
sodium to enter the muscle fiber. As always, if the depolarization reaches the
threshold, an action potential is formed. It spreads across the plasmalemma
into the T-tubules, inititating muscle fiber contraction. As in the neuron, the
plasmalemma, once depolarized, must undergo repolarization. During the period
of repolarization, the sodium gates are closed and the potassium gates are
open; thus, like the neuron, the muscle fiber is unable to respond to any
further stimulation. This period is reffered to as the refractory period. Once
the electrical conditions of the muscle fiber are restored to resting levels,
the fiber can respond to another stimulus. Thus, the refractory period limits
the motor unit’s firing frequency.
Now we know how the impulse is transmitted
between two cells. But to understand what happens once the impulse is
transmitted, we must first examine the chemical signals that accomplish
transmission.
Neurotransmitters
More than 50 neurotransmitters have been
positively identified or are suspected as potential candidates. These can be
cathegorized as either (a) small-molecule, rapid-acting neurotransmitters
or (b) neuropeptide, slow-acting neurotransmitters. The small-molecule,
rapid-acting transmitters, which are responsible for most neural transmissions,
are our main concern.
Acetylholine and norepinephrine are the two
major neurotransmitters involved in regulating our physiological responses to
exercise. Acetylholine is the
primary neurotransmitter for the motor neurons that innervate skeletal muscle
and for most parasympathetic neurons. It is generally an excitatory
neurotransmitter, but it can have inhibitory effects at some parasympathetic
nerve endings, such as in the heart. Norepinephrine
is the neurotransmitter for the most sympathetic neurons, and it too can be
either excitatory or inhibitory, depending on the receptors involved.
Once the neurotransmitters binds to the
post-synaptic receptor, the nerve impulse has been successfully transmitted.
The neurotransmitter is then either degraded by enzymes, actively transported
back into the presynaptic terminals for reuse, or diffused away from the
synapse.
Synapse
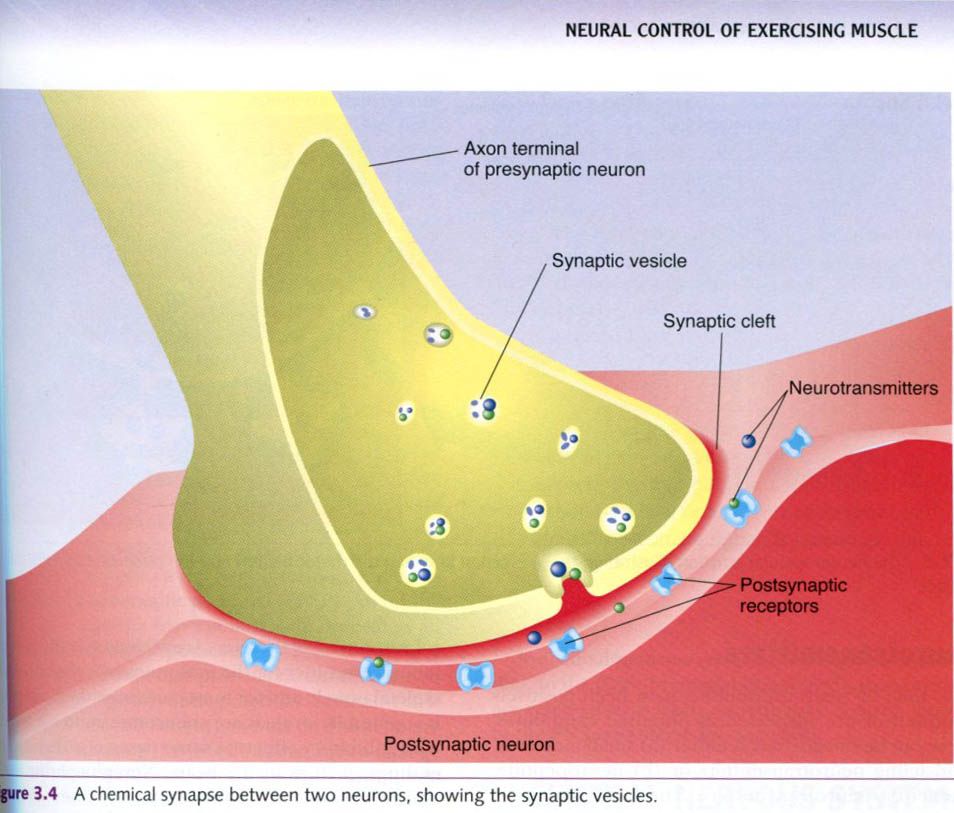
Neurons communicate with each other across
synapses. A synapse is the site
of action potential transmission from one neuron to another. There are both
chemical and mechanical synapses, but the most common type is the
chemical synapse, which is our focus. As seen in the figure up, a synapse
between two neurons includes:
- The axon terminals of the
neuron sending the action potential;
- Receptors on the neuron
receiving the action potential, and
- The space between these structures.
The neuron sending the action potential
across the synapse is called the presynaptic
neuron, so axon terminals are presynaptic terminals. Similarly, the
neuron receiving the action potential on the opposite side of the synapse
is called the postsynaptic receptors.
The axon terminals and postsynaptic receptors are not physically in contact
with each other. A narrow gap, the synaptic
cleft, separates them.
The action potential can be transmitted
across a synapse in only one direction: from the axon terminals of the
presynaptic neuron to the postsynaptic receptors, usually on the dendrites, of
the postsynaptic neuron. Impulses also can go directly to receptors on the
cell body: about 5% to 20% of the axon terminals are adjacent to the cell body
instead of the dendrites. Why can the action potential go in only one
direction?
The presynaptic terminals of the axon contain a
large number of saclike structures, called synaptic
vesicles. These sacs contain neurotransmitter chemicals. When the impulse
reaches the presynaptic axon terminals, the synaptic vesicles respond by
dumping their chemicals into the synaptic cleft. These neurotransmitters then
diffuse across the synaptic cleft to the postsynaptic neuron’s receptors. The
postsynaptic receptors bind the neurotransmitter once it diffuses across the
synaptic cleft. When this binding occurs, the impulse has been transmitted
successfully to the next neuron and can be transmitted onward.
30. 5. 2012.
Nerve impulse
Resting membrane potential
The cell membrane of a neuron at rest has a
negative electrical potential of about -70mV. This means that if one were to
insert a voltmeter probe inside the cell, the electrical charges found there
and the charges found outside the cell would differ by 70mV, and the inside
would be negative relative to the outside. This electrical potential difference
is known as the resting membrane
potential(RMP). It is caused by a separation of charges across the membrane
differ, the membrane is said to be polarized.
The neuron has a high concentration of potassium ions(K+) on the
inside of the membrane and a high concentration of sodium ions(Na+)
on the outside. The imbalance in the number of ions inside and outside the cell
causes the RMP. The imbalance in the number of ions inside and outside the cell
causes the RMP. This imbalance is maintained in two ways. First, the cell membrane
is much more permeable to K+ than to Na+, so the K+
can move more freely. Because ions tend to move to establish equilibrium, some
of the K+ will move to an area where it is less concentrated,
outside the cell. The Na+ cannot move to the inside as easily.
Second, sodium-potassium pumps in
the neuron membrane, which contain Na+-K+ adenosine
triphosphatase(ATPase), maintain the imbalance on each side of the membrane by
actively transporting potassium ions
in the sodium ions out. The sodium-potassium pump moves three Na+
out of the cell for each two K+ it brings in. The end result is that
more positively charged ions are outside the cell than inside, creating the
potential difference across the membrane. Maintenance of a constant RMP of
about -70mV is primarily a function of the sodium-potassium pump.
Depolarization and hyperpolarization
If the inside of the cell becomes less negative
relative to the outside, the potential difference across the membrane
decreases. The membrane will be less polarized. When this happens, the membrane
is said to be depolarized. Thus, depolarization
occurs any time the charge difference becomes less than the RMP of -70mV,
moving closer to zero. This typically results from a change in the membrane’s
Na+ permeability.
The opposite can also occur. If the charge
difference across the membrane increases, moving from the RMP to an even more
negative value, then the membrane becomes more polarized. This is known as hyperpolarization. Changes in the
membrane potential are actually signals used to receive, transmit, and
integrate information within and between cells. These signals are of two types,
graded potentials and action
potentials. Both are electrical currents created by the movement of
ions.
Graded potentials
Graded
potentials are localized
changes in the membrane potential. These changes can be either depolarizations
or hyperpolarizations. The membrane contains ion channels with ion gates that
act as doorways into and out of the neuron. These gates are usually closed, preventing
ion flow; but they open with stimulation; allowing ions to move from the
outside to the inside or vice versa. This ion flow alters the charge
separation, changing the polarization of the membrane.
Graded potentials are triggered by a change in
the neuron’s local environment. Depending on the location and type of neuron
involved, the ion gates may open in response to the transmission of an impulse
from another neuron or in response to sensory stimuli such as changes in
chemical concentrations, temperature, or pressure.
Recall that most neuron receptors are located
on the dendrites( although some are on the cell body), yet the impulse is
always transmitted from the axon terminals at the opposite end of the cell. For
a neuron to transmit an impulse, the impulse must travel almost the entire
length of the neuron. Although a graded potential may result in depolarization
of the entire cell membrane, it is usually just a local event such that the
depolarization does not spread very far along the neuron. To travel the full
distance, an impulse must generate an action potential.
Action potentials
An action
potential is a rapid and substantial depolarization of the neuron’s
membrane. It usually lasts about 1ms. Typically, the membrane potential changes
from the RMP of about -70mV to a value of about +30mV and then rapidly returns
to its resting value. This is illustrated in the picture below. How does this
marked change in membrane potential occur?
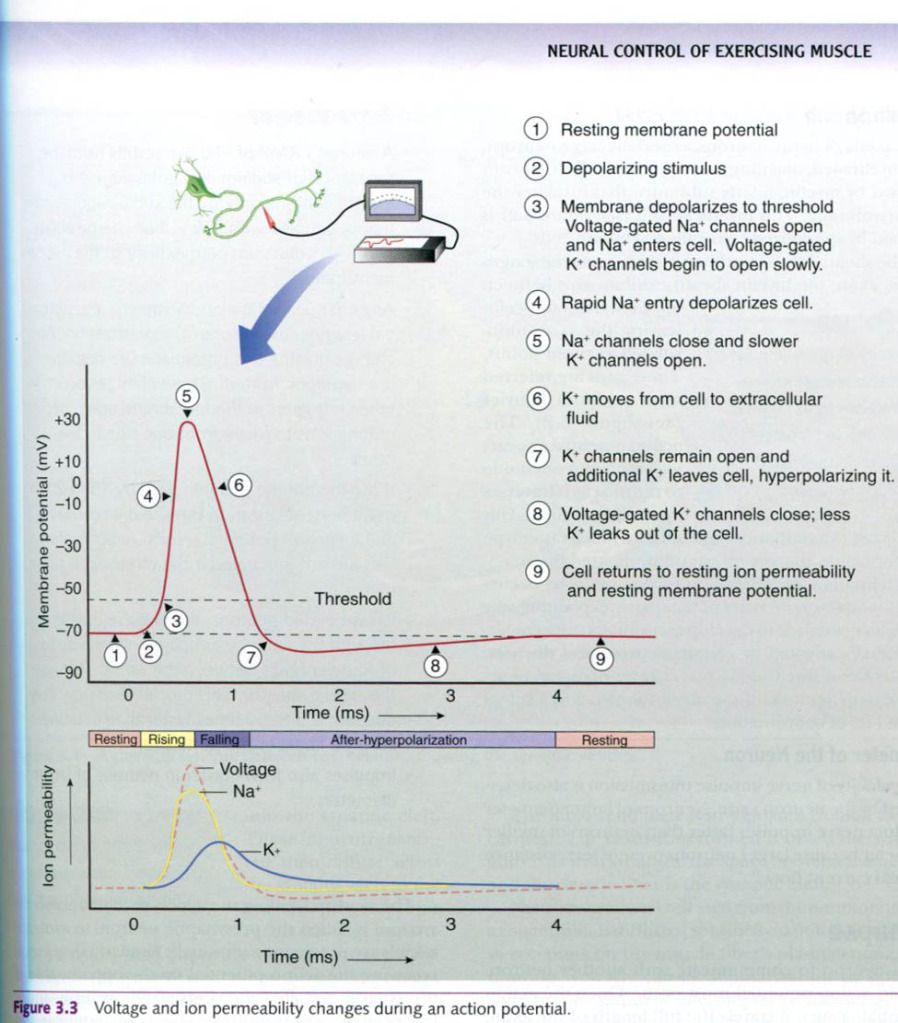
All action potentials begin as graded
potentials. When enough stimulation occurs to cause a depolarization of at
least 15 to 20mV, an action potential results. In other words, if the membrane
depolarizes from the RMP of -70mV to a value of -50 to -55mV, the cell will
experience an action potential. The membrane voltage at which a graded
potential becomes an action potential is called the depolarization threshold.
Any depolarization that does not attain the threshold will not result in an
action potential. For example, if the membrane potential changes from the RMP of
-70mV to -60mV, the change is only 10mV and does not reach the threshold; thus,
no action potential occurs. But any time depolarization reaches or exceeds the
threshold, an action potential will result. This is the all-or-none principle.
When a given segment of an axon is generating
an action potential and its sodium gates are open, it is unable to respond to
another stimulus. This is reffered to as the absolute refractory period. When the sodium gates are
closed, the potassium gates are open, and repolarizing is occurring, the
segment of the axon can then respond to a new stimulus, but the stimulus must
be of substantially greater magnitude to evoke an action potential. This is
reffered to as the relative refractory
period.
Propagation of the action potential
Now that we understand how a neural impulse, in
the form of an action potential, is generated, we can look at how the impulse
is propagated, or how it travels through the neuron. Two characteristics of the
neuron become particularly important when we consider how quickly an impulse
can pass through the axon: myelination
and diameter.
Myelin
sheath
The axons of many neurons, especially large
neurons, are myelinated, meaning they are covered with a sheath formed by
myelin, a fatty substance that insulates the cell membrane. This myelin sheath is formed by specialized
cells called Schwann cells.
The sheath is not continuous. As it spans the
length of the axon, the myelin sheath exhibits gaps between adjacent Schwann
cells, leaving the axon uninsulated at those points. These gaps are reffered to
as nodes of Ranvier. The action
potential appears to jump from one node to the next as it transverses a
myelinated fiber. This is reffered to as salutatory
conduction, a much faster type of conduction than occurs in unmyelinated
fibers.
Myelination of peripheral motor neurons occurs
over the first several years of life, partly explaning why children need time
to develop coordinated movement. Individuals affected by certain neurological
diseases, such as MS as discussed in our chapter opening, experience
degeneration of the myelin sheath and a subsequent loss of coordination.
Diameter
of the neuron
The velocity of nerve impulse transmission is
also determined by the neuron’s size. Neurons of larger diameter conduct nerve
impulses faster than neurons of smaller diameter because larger neurons present
less resistance to local current flow.
Overview of the nervous system
Structure and function of the nervous system
The neuron is the structural unit of the
nervous system. We first review the anatomy of the neuron and then look at how
it functions – allowing electrical impulses to be transmitted throughout the body.
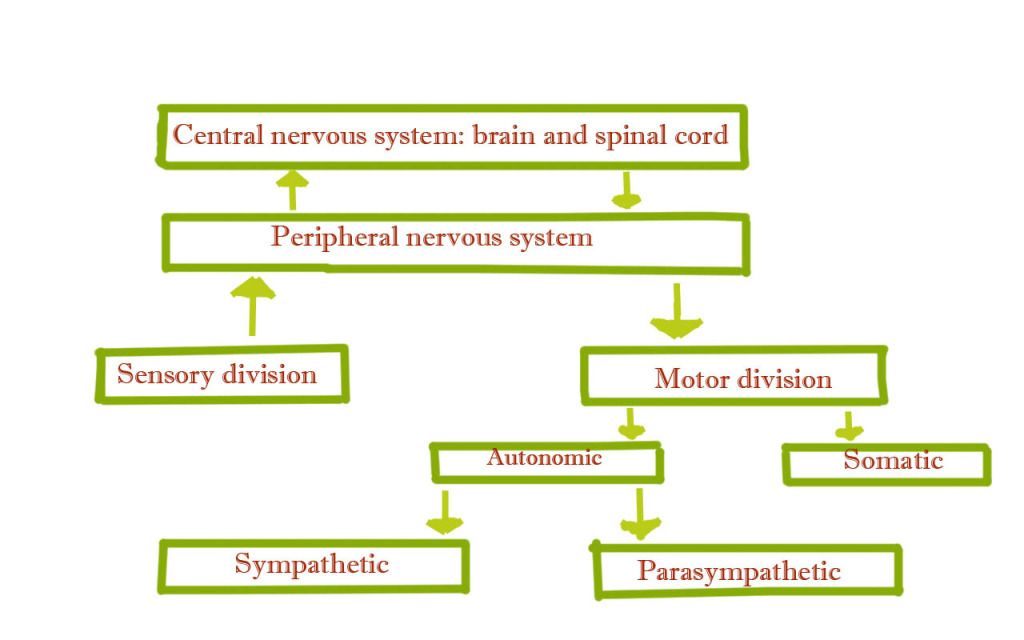
Neuron
Individual nerve fibers(nerve cells), depicted
in the figure below, are called neurons. A typical neuron is composed of
three regions:
- The cell body, or soma
- The dendrites
- The axon
The cell body contains the nucleus.
Radiating out from the cell body are the cell processes; the dendrites and the
axon. On the side toward the axon, the cell body tapers into a cone-shaped
region region known as the axon hillock.
The axon hillock has an important role in impulse conduction.
Most neurons contain many dendrites. These are
the neuron’s receivers. Most impulses, or action potentials, coming into the
neuron from sensory stimuli or from adjacent neurons typically enter the neuron
via the dendrites. These processes then carry the impulses toward the cell
body.
In contrast, most neurons have only one axon.
The axon is the neuron’s transmitter and conducts impulses away from the cell
body. Near its end, an axon splits into numerous end branches. The tips of these branches are dilated into tiny
bulbs known as axon terminals or
synaptic knobs. These terminals or knobs house numerous vesicles(sacs) filled
with chemicals known as neurotransmitters
that are used for communication between a neuron and another cell. The
structure of the neuron allows nerve impulses to enter the neuron through the
dendrites, and to a lesser extent through the cell body, and to travel through
the cell body and axon hillock, down the axon, and out through the end branches
to the axon terminals. We next explain in more detail how this happens,
including how these impulses travel from one neuron to another and from a motor
neuron to muscle fibers.

29. 5. 2012.
Hormonal regulation of fluid and electrolyte balance during exercise
The endocrine system plays a major role in
monitoring fluid levels and correcting imbalances, along with regulating
electrolyte balance, especially that of sodium. The two major hormones involved
in this regulation are antidiuretic hormone released from the posterior
pituitary and aldosterone, a mineralocorticoid released from the adrenal
cortex. The kidneys are the primary target organ for both of these hormones.
Posterior pituitary
The pituitary’s posterior lobe is an outgrowth
of neural tissue form the hypothalamus. For this reason, it is also reffered to
as the neurohypophysis. It secretes two hormones; antidiuretic hormone(ADH; also called vasopressin or arginine
vasopressin) and oxytocin. Both of these hormones are actually produced in the
hypothalamus. They travel through the neural tissue and are stored in vesicles
within nerve endings in the posterior pituitary. These hormones are released
into capillaries as needed in response to neural impulses from the
hypothalamus.
Of the two posterior pituitary hormones, only
ADH is known to play an important role during exercise. Antidiuretic hormone
promotes water conservation by increasing the water permeability of the
kidneys’ collecting ducts. As a result, less water is excreted in the urine,
creating an “antidiuresis”.
Muscular activity and sweating cause
electrolytes to become concentrated in the blood plasma. This is called hemoconcentration, and it increases the
plasma osmolality(the ionic
concentration of dissolved substances in the plasma). This is the primary
physiological stimulus for ADH release. The increased osmolality is sensed by
osmoreceptors in the hypothalamus. A second and related stimulus for ADH
release is a low plasma volume. In response to either stimuli, the hypothalamus
sends neural impulses to the posterior pituitary, stimulating ADH release. The
ADH enters the blood, travels to the kidneys, and promotes water retention in
an effort to dilute the plasma electrolyte concentration back to normal levels.
This hormone’s role in conserving body water minimizes the extent of water loss
and therefore the risk of severe dehydration during periods of heavy sweating
and hard exercise. Picture below illustrates this process.
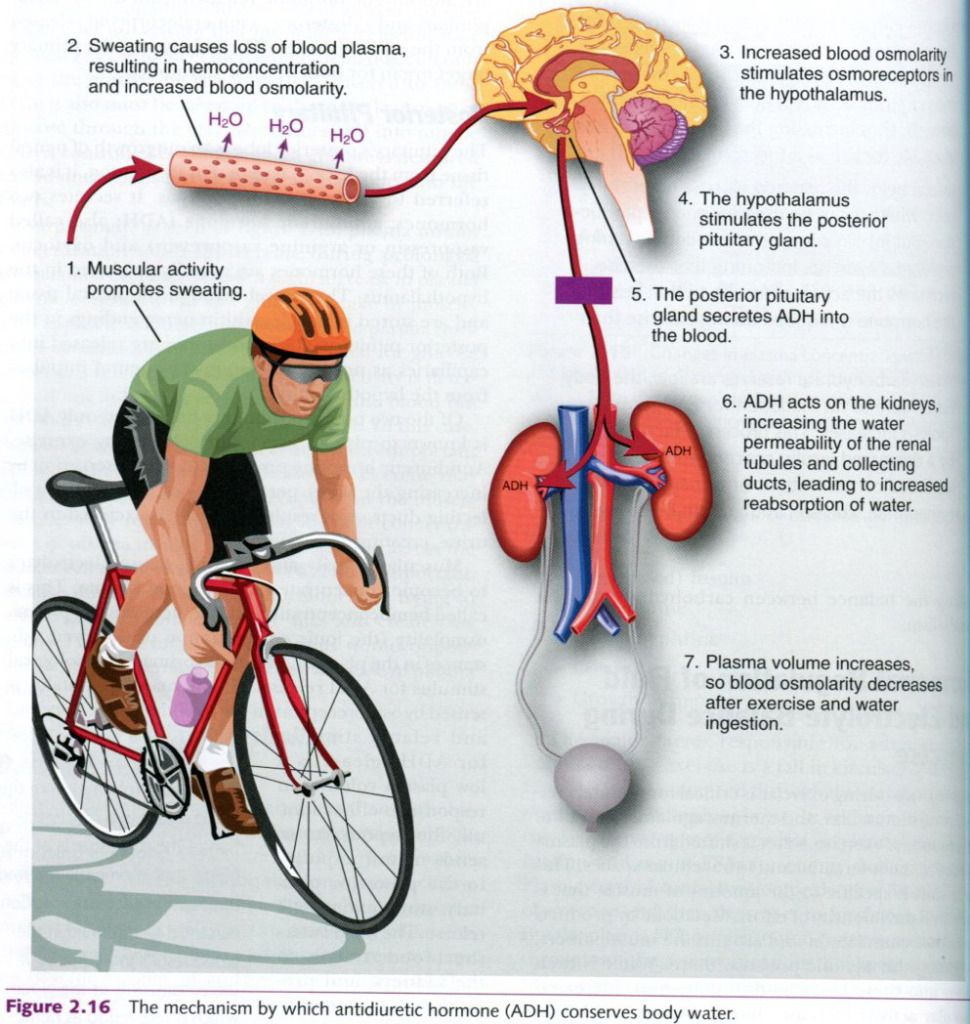
Adrenal cortex revisited
The mineralocorticoids,
secreted from the adrenal cortex, maintain electrolyte balance in the
extracellular fluids, especially that of sodium(Na+) and potassium(K+). Aldosterone is the major
mineralocorticoid, responsible for at least 95% of all mineralocorticoid
activity. It works primarily by promoting renal reabsorption of sodium, thus
causing the body to retain sodium. When sodium is retained, so is water: thus,
aldosterone, like ADH, results in water retention. Sodium retention also
enhances potassium excretion, so
aldosterone plays a role in potassium balance as well. For these reasons, aldosterone
secretion is stimulated by many factors, including decreased plasma sodium,
decreased blood volume, decreased blood pressure, and increased plasma
potassium concentration.
Kidneys
Although the kidneys are not typically
considered major endocrine organs, they release a hormone called erythropoetine. Erythropoetin(EPO)
regulates red blood cell(erythrocyte) production by stimulating bone marrow
cells. The red blood cells are essential for transporting oxygen to the
tissues and removing carbon dioxide, so this hormone is extremely important in
our adaptation to training and altitude. The kidneys also release renin, a
hormone and enzyme involved in blood pressure control and fluid and electrolyte
balance.
The kidneys have a strong regulatory influence
on blood pressure that also allows them to regulate fluid balance. Plasma
volume is a major determinant of blood pressure: when plasma volume decreases,
so does blood pressure. Blood pressure is monitored by specialized cells within
the kidneys. During exercise, these cells can be stimulated by decreased blood
pressure, decreased blood flow to the kidneys through increased sympathetic nervous
activity accompanying exercise, or direct stimulation from the sympathetic
nerves.
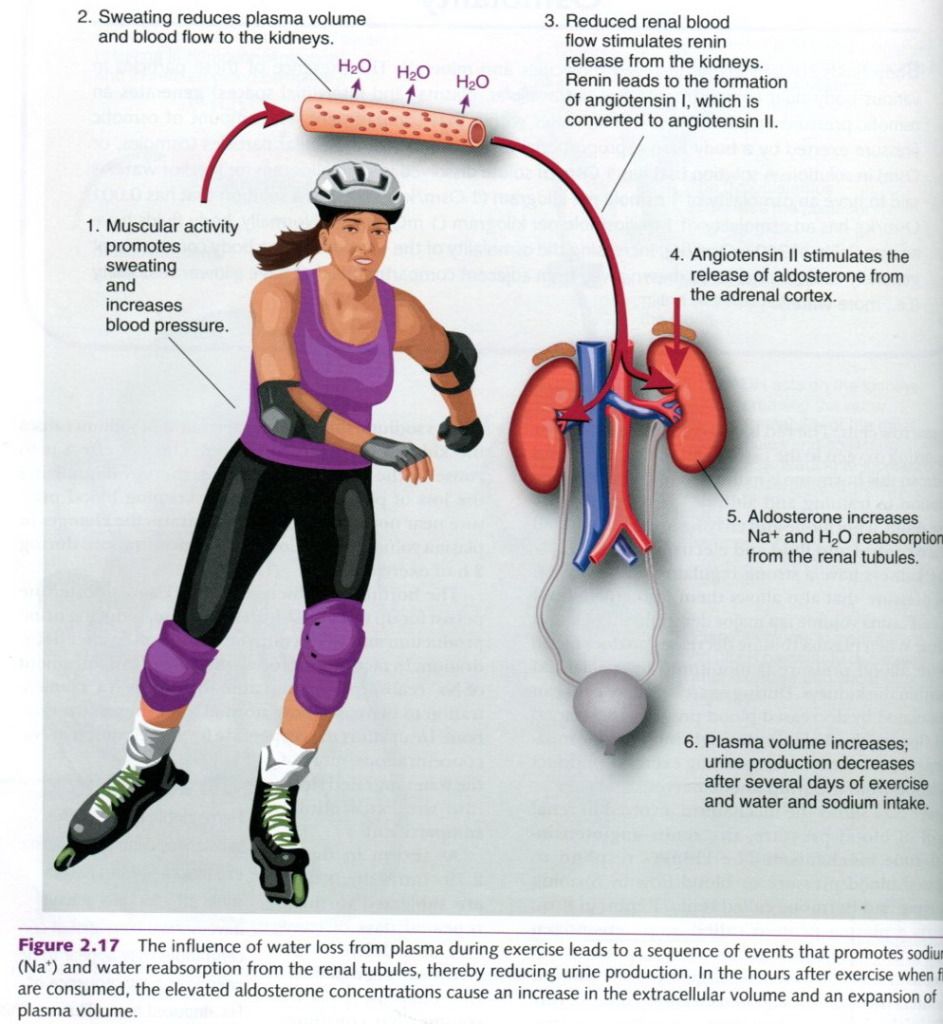
Figure shows the mechanism involved in renal
control of blood pressure, the
renin-angiotensin-aldosterone mechanism. The kidneys respond to decreased
blood pressure or blood flow by forming an enzyme and hormone called renin. Renin, in turn, converts a
plasma protein called angiotensinogen into an active form called angiotensin I.
In the blood, angiotensin I is converted to angiotensin II when it encounters
the enzyme angiotensin converting
enzyme(ACE) in the lungs. Angiotensin converting enzyme inhibitors are a
class of drugs used in the treatment of high blood pressure. They lower blood
pressure by blocking, or inhibiting, the conversion of angiotensin I to angiotensin
II. Angiotensin II acts in two ways. First, it is a potent constrictor of blood
vessels. Through this action, peripheral resistance increases, which raises the
blood pressure. The second job of angiotensin II is to trigger aldosterone
release from the adrenal cortex.
Recall that aldosterone’s primary action is to
promote sodium reabsorption in the kidneys. Because water follows sodium, this
renal conservation of sodium causes the kidneys to also retain water. The net
effect is to conserve body’s fluid content, thereby minimizing the loss of
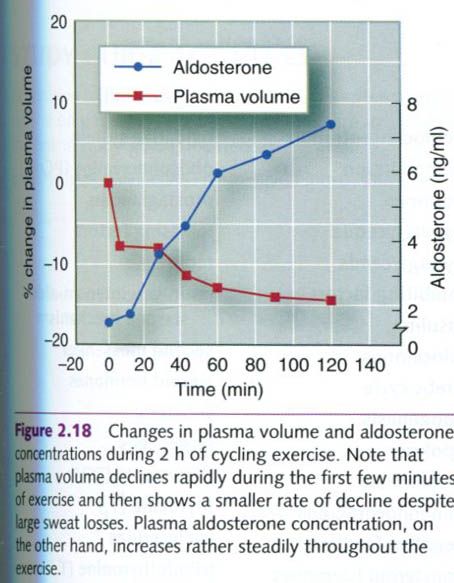
plasma volume while keeping blood pressure near normal. Figure below
illustrates the changes in plasma volume and aldosterone concentrations during
2h of exercise.
The hormonal influences of ADH and aldosterone
persist for up to 12 to 48h after exercise, reducing urine production and
protecting the body from further dehydration. In fact, aldosterone’s prolonged
enhancement of Na+ reabsorption may cause the body’s Na+
concentration to increase above normal following an exercise bout.
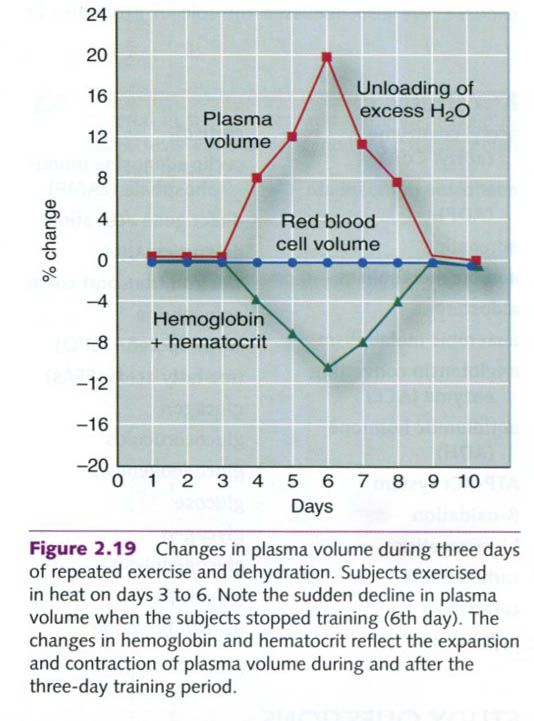
As shown in the figure on the right side, individuals who
are subjected to three repeated days of exercise and dehydration show a
significant increase in plasma volume that continues to increase throughout the
period of activity. This increase in plasma volume appears to parallel the
body’s retention of dietary Na+. When the daily bouts of activity
are terminated, the excess Na+ and water are excreted in urine.
Most athletes involved in heavy training have
an expanded plasma volume, which dilutes various blood constituents. The actual
amount of proteins and electrolyte(solutes) within the blood remains unaltered,
but the substances are dispersed throughout a greater volume of water(plasma),
so they are diluted and their concentration decreases. This phenomenon is
called hemodilution.
“Physiology of sport and exercise”, fourth edition; Jack H. Wilmore, David L. Costill, W. Larry Kenney
“Physiology of sport and exercise”, fourth edition; Jack H. Wilmore, David L. Costill, W. Larry Kenney
Hormonal regulation of metabolism during exercise
Regulation of glucose metabolism during exercise
The heightened energy demands of exercise
require that more glucose be made available to the muscles. Recall that glucose
is stored in the body as glycogen,
primarily in the muscles and the liver. Glucose must be freed from its storage
form of glycogen, so glycogenolysis must increase. Glucose freed from the liver
enters the blood to circulate throughout the body, allowing it access to active
tissues. Plasma glucose concentration also can be increased through gluconeogenesis.
Regulation of plasma glucose concentration
Four hormones work to increase the amount of
circulating plasma glucose:
- Glucagon
- Epinephrine
- Norepinephrine
- Cortisol
The plasma glucose concentration during
exercise depends on a balance between glucose uptake by exercising muscles and
its release by the liver. At rest, glucose release from the liver is
facilitated by glucagon, which promotes both liver glycogen breakdown and
glucose formation from amino acids. During exercise, glucagon secretion
increases. Muscular activity also increases the rate of catecholamine release
from the adrenal medulla, and these
hormones(epinephrine and norepinephrine) work with glucagons to further
increase glycogenolysis. Cortisol concentrations also increase during exercise.
Cortisol increases protein
catabolism, freeing amino acids to be used within the liver for gluconeogenesis. Thus, all four of these
hormones can increase plasma glucose by enhancing the processes of glycogenolysis(breakdown
of glycogen) and gluconeogenesis(making glucose from other substrates). In
addition to the effects of the four major glucose-controlling hormones, growth hormone increases mobilization of
FFAs and decreases cellular uptake of glucose, so less glucose is used by the
cells(more remains in circulation); and the thyroid hormones promote glucose
catabolism and fat metabolism.
The amount of glucose released by the liver
depends on exercise intensity and the duration. As intensity increases, so does
the rate of catecholamine release. This can cause the liver to release more
glucose than is being taken up by the active muscles. Consequently, during or
shortly after an explosive, short-term sprint bout, blood glucose
concentrations may be 40% to 50% above the resting level, illustrating that the
glucose release by the liver is greater than the uptake by the muscles.
The greater the exercise intensity, the greater
the catecholamine release, and thus the glycogenolysis rate is significantly
increased. This process occurs not only in the liver but also in the muscle.
The glucose released from the liver enters the blood to become available to the
muscle. But the muscle has a more readily available source of glucose: its own
glycogen. The muscle uses its own glycogen stores before using the plasma
glucose during explosive, short-term exercise. Glucose released from the liver
is not used as readily, so it remains in the circulation, elevating the plasma
glucose. Following exercise, plasma glucose concentrations decrease as the
glucose enters the muscle to replenish the depleted muscle glycogen stores.
During exercise bouts that last for several
hours, however, the rate of liver glucose release more closely matches the
muscles’ needs, keeping plasma glucose at or only slightly above the resting
concentrations. As muscle uptake of glucose increases, the liver’s rate of
glucose release also increases. In most cases, plasma glucose does not begin to
decline until late in the activity as liver glycogen stores become depleted, at
which time glucagons concentrations increase significantly. Glucagon and cortisol together enhance gluconeogenesis,
providing more fuel.
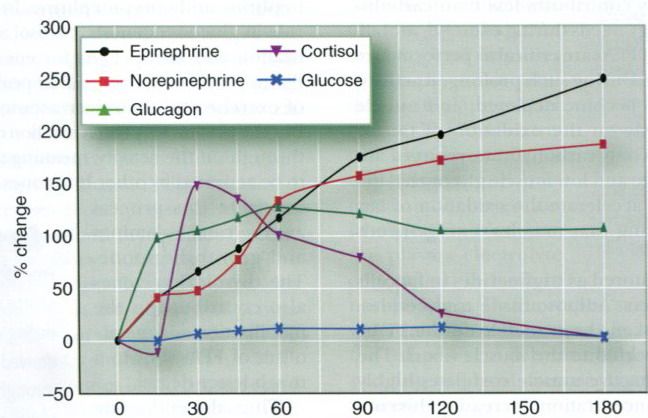
Figure below illustrates the changes in plasma
concentrations of epinephrine, norepinephrine, glucagon, cortisol, and glucose
during 3h of cycling. Although the hormonal regulation of glucose remains
intact throughout such longterm activities, the liver’s glycogen supply may
become critically low. As a result, the liver’s rate of glucose release may be
unable to keep pace with the muscles’ rate of glucose uptake. Under this
condition, plasma glucose may decline despite strong hormonal stimulation.
Glucose ingestion during the activity can play a major role in maintaining
plasma glucose concentrations.
Glucose uptake by muscle
Merely releasing sufficient amounts of glucose
into the blood does not ensure that the muscle cells will have enough glucose
to meet their energy demands. Not only must the glucose be released and
delivered to these cells; it also must be taken up by the cells. Transport of
glucose through the cell membranes and into muscle glucose through the cell
membranes and into muscle cells in controlled by insulin. Once glucose is delivered to the muscle, insulin
facilitates its transport into the fibers.
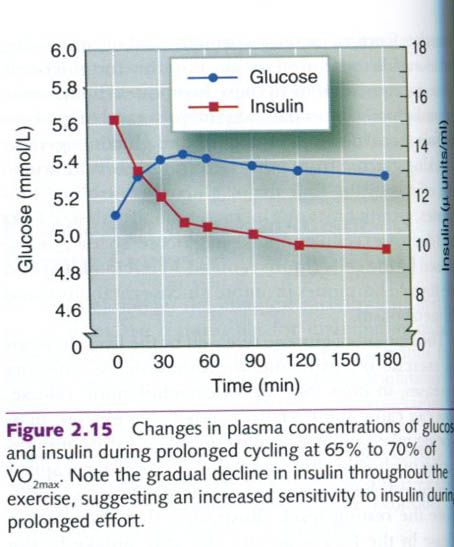
Surprisingly, as seen in figure below, plasma
insulin concentration tends to decrease during prolonged submaximal exercise,
despite a slight increase in plasma glucose concentration and glucose uptake by
muscle. This apparent contradiction between the plasma insulin concentrations
and the muscles’ need for glucose serves as a reminder that a hormone’s
activity is determined not only by its concentration in the blood but also by a
cell’s sensitivity to that hormone. Exercise may enhance insulin’s binding to
receptors on the muscle fiber, thereby reducing the need for high
concentrations of plasma insulin to transport glucose across the muscle cell
membrane into the cell. This is important, because during exercise four
hormones are trying to release glucose from its storage sites and create new
glucose. High insulin concentrations would oppose their action, preventing this
needed increase in plasma glucose supply.
Regulation of fat metabolism during exercise
Although fat generally contributes less than
carbohydrate to muscles’ energy needs during exercise, mobilization and
oxidation of FFAs are critical to performance in endurance exercise. During
such prolonged activity, carbohydrate reserves become depleted, and muscle must
rely more heavily on the oxidation of fat for energy production. When
carbohydrate reserves are low(low plasma glucose and low muscle glycogen), the
endocrine system can accelerate the oxidation of fats(lypolysis), thus ensuring
that muscles’ energy needs can be met.
Free fatty acids are stored as triglycerides in
fat cells and inside muscle fibers. Adipose tissue triglycerides, however, must
be broken down to release the FFAs, which are then transported to the muscle
fibers. The rate of FFA uptake by active muscle correlates highly with the
plasma FFA concentration. Increasing this concentration would increase cellular
uptake of the FFA. The rate of triglyceride breakdown may determine, in part,
the rate at which muscles use fat as a fuel source during exercise.
The rate of lypolysis is controlled by at least
five hormones:
- (decreased) Insulin
- Epinephrine
- Norepinephrine
- Cortisol
- Growth hormone
The major factor responsible for adipose tissue
lypolysis during exercise is a fall in circulating insulin. Lipolysis is also
enhanced through the elevation of epinephrine and norepinephrine. In addition
to having a role in gluconeogenesis,
cortisol accelerates the mobilization and use of FFAs for energy during
exercise. Plasma cortisol concentration peaks after 30 to 45 min of exercise
and then decreases to near-normal levels. But the plasma FFA concentration
continues to increase throughout the activity, meaning that lipase continues to
be activated by other hormones. The hormones that continue this process are the
catecholamines and growth hormone. The thyroid hormones also contribute to the
mobilization and metabolism of FFAs, but to a much lesser degree.
Thus, the endocrine system plays a critical
role in regulating ATP production during exercise as well as controlling the
balance between carbohydrate and fat metabolism.
“Physiology of sport and exercise”, fourth
edition; Jack H. Wilmore, David L. Costill, W. Larry Kenney
28. 5. 2012.
Hormonal response to acute exercise and change in response with exercise training
Schema will be following: HORMONE – RESPONSE TO ACUTE EXERCISE(UNTRAINED) – EFFECT OF EXERCISE
TRAINING
Anterior
pituitary gland
Growth
hormone(GH) – increases with
increasing rates of work – attenuated response at the same rate of work
Thyrotropin(TSH) – increases with increasing rates of work – no
known effect
Adrenocorticotropin(ACTH) – increases with increasing rates of work and
duration – attenuated response at same rate of work
Prolactin – increases with exercise – no known effect
Follicle-stimulating
hormone(FSH) – small or no
change – no known effect
Luteinizing
hormone(LH) – small or no
change – no known effect
Posterior
pituitary
Antidiuretic
hormone(ADH or vasopressin) –
increases with increasing rates of work – attenuated response at same rate of
work
Oxytocin – unknown – unknown
Thyroid
Thyroxine(T4)
and triiodothyronine(T3) – free T3 and T4 increase with increasing rates
of work – increased turnover of T3 and T4 at same rate of
work
Calcitonin – unknown – unknown
Parathyroid
Parathyroid
hormone(PTH or parathormone) –
increases with prolonged exercise – unknown
Adrenal
medulla
Epinephrine – increases with increasing rates of work,
starting at about 75% of VO2max – attenuated response at same rate
of work
Norepinephrine – increases with increasing rates of work,
starting at about 50% of VO2max – attenuated response at same rate
of work
Adrenal
cortex
Aldosterone
– increases with increasing
rates of work – unchanged
Cortisol
– increases only at high rates
of work – slightly higher values
Pancreas
Insulin – decreases with increasing rates of work –
attenuated response at same rate of work
Glucagon
– increases with increasing
rates of work – attenuated response at same rate of work
Kidney
Renin – increases with increasing rates of work –
unchanged
Erythropoetin(EPO) – unknown – unchanged
Testes
Testosterone – small increases with exercise – resting
level decreases in male runners
Ovaries
Estrogens
and progesterone – small
increases with exercise – resting levels might be decreased in highly trained
women
“Physiology of sport and exercise”, fourth edition; Jack H. Wilmore, David L. Costill, W. Larry Kenney
“Physiology of sport and exercise”, fourth edition; Jack H. Wilmore, David L. Costill, W. Larry Kenney
Endocrine glands and their hormones – overview
The pituitary gland is a marble-sized gland at
the base of the brain. The secretory action of the pituitary is controlled by
either neural mechanisms or hormones secreted by the hypothalamus. Therefore,
the pituitary gland can be thought of as the relay between central nervous
system control centers and peripheral endocrine glands.
The pituitary gland is composed of three lobes:
anterior, intermediate and posterior. The intermediate lobe is very small and is thought to play little or
no role in humans, but both the posterior and anterior lobes have major
endocrine functions. The anterior pituitary has a major role in fluid and
electrolyte balance.
The anterior pituitary, also called the adenohypophysis,
secretes six hormones in response to releasing
factors and inhibiting factors(hormones)
secreted by the hypothalamus. Communication between the hypothalamus and the
anterior lobe of the pituitary occurs through a specialized circulatory system
that transports the releasing and inhibiting factor from the hypothalamus to
the anterior pituitary. The major functions of each of the anterior pituitary
hormones, along with their releasing and inhibiting factors are discussed here. Exercise appears to be a strong
stimulant to the hypothalamus because exercise increases the release rate of
all anterior pituitary hormones.
Of the six anterior pituitary hormones, four
are tropic hormones, meaning they affect the functioning of other endocrine
glands. The exceptions are growth hormone and prolactin. Growth hormone is a potent anabolic agent(a substance that promotes
constructive metabolism). It promotes muscle growth and hypertrophy by
facilitating amino acid transport into the cells. In addition, growth hormone
directly stimulates fat metabolism(lypolysis) by increasing the synthesis of
enzymes involved in this process. Growth hormone concentrations are elevated
during aerobic exercise, apparently in proportion to the exercise intensity,
and typically remain elevated for some time after exercise.
Thyroid gland
The thyroid gland is located along the midline
of the neck, immediately below the larynx. It secretes two important nonsteroid
hormones, triiodothyronine(T3)
and thyroxine(T4), which regulate metabolism in general, and an
additional hormone, calcitonin, which assists in regulating calcium metabolism.
The two metabolic thyroid hormones share
similar functions. Triiodothyronine and thyroxine increase the metabolic rate
of almost all tissues and can increase the body’s basal metabolic rate by as
much as 60% to 100%. These hormones also:
- Increase protin
synthesis(also enzyme synthesis);
- Increase the size and number
of mitochondria in most cells;
- Promote rapid cellular
uptake of glucose;
- Enhance glycolysis and gluconeogenesis;
- Enhance lipid mobilization,
increasing FFA availability for oxidation.
Release of
thyrotropin(thyroid-stimulating hormone, or TSH) from the anterior pituitary increases during exercise.
Thyroid-stimulating hormone controls the release of triiodothyronine and
thyroxine, so the exercise-induced increase in TSH would be expected to
stimulate the thyroid gland. Exercise does increase plasma thyroxine
concentrations, but a delay occurs between the increase in TSH concentrations
during exercise and the increase in TSH concentrations during exercise and the
increase in plasma thyroxine concentration. Furthermore, during prolonged
submaximal exercise, thyroxine concentrations remain relatively constant after
a sharp initial increase as exercise begins, and triiodothyronine
concentrations tend to decrease.
Adrenal glands
The adrenal glands are situated directly atop
each kidney and are composed of the inner adrenal medulla and the outer adrenal
cortex. The hormones secreted by these two parts are quite different, so we
consider them separately. The adrenal medulla produces and releases two
hormones, epinephrine and norepinephrine, which are collectively
referred to as catecholamines.
Because of its origin in the adrenal gland, a synonym for epinephrine is adrenaline. When the adrenal medulla is
stimulated by the sympathetic nervous system, approximately 80% of its
secretion is epinephrine and 20% is norepinephrine, although these percentages
vary with different physiological conditions. The catecholamines have powerful
effects similar to those of the sympathetic nervous system. Recall that these
same catecholamines function as neurotransmitters in the sympathetic nervous
system: however, the hormones’ effect last longer because these substances are
removed from the blood relatively slowly compared to the quick reuptake and
degradation of the neurotransmitters. These two hormones prepare a person for
immediate action, often called the “fight-or-flight response”.
Although some of the specific actions of these
two hormones differ, the two work together. Their combined effects include:
- Increased heart rate and
force of concentration;
- Increased metabolic rate;
- Increased
glycogenolysis(breakdown of glycogen to glucose) in the liver and muscle;
- Increased release of glucose
and FFAs into the blood;
- Redistribution of blood to
the skeletal muscles(through vasodilatation of vessels supplying skeletal
muscles and vasoconstriction of vessels to the skin and viscera);
- Increased blood pressure;
- Increased respiration.
Release of epinephrine and norepinephrine is
affected by a wide variety of factors, including changes in body position,
psychological stress, and exercise. Plasma concentrations of these hormones
increase as individuals gradually increase their exercise intensity. Plasma
norepinephrine concentrations increase markedly at work rates above 50% of VO2max,
but epinephrine concentrations do not increase significantly until the exercise
intensity exceeds 60% to 70% of VO2max. During long-duration
steady-state activity of moderate intensity, blood concentrations of both
hormones increase. When the exercise bout ends, epinephrine returns to resting
concentrations within only a few minutes of recovery, but norepinephrine can
remain elevated for several hours.
The adrenal cortex secretes more than 30
different steroid hormones, referred to as corticosteroids. These generally are
classified into three major types: mineralocorticoids, glucocorticoids, and
gonadocorticoids(sex hormones).
The glucocorticoids
are essential components in the ability to adapt to external changes and
stress. They also help maintain fairly consistent plasma glucose concentrations
even when we go for long periods without ingesting food. Cortisol, also known as hydrocortisone, is the major
corticosteroid. It is responsible for about 95% of all glucocorticoid activity
in the body. Cortisol:
- Stimulates gluconeogenesis to ensure an
adequate fuel supply;
- Increases mobilization of
FFAs, making them more available as an energy source;
- Decreases glucose
utilization, sparing it for the brain;
- Stimulates protein
catabolism to release amino acids for use in repair, enzyme synthesis, and
energy production;
- Acts as an anti-inflammatory
agent;
- Depresses immune reactions;
- Increases the
vasoconstriction caused by epinephrine.
Pancreas
The pancreas is located behind and slightly
below the stomach. Its two major hormones are insulin and glucagon. The balance
of these two opposing hormones provides the major control of plasma glucose
concentrations. When plasma glucose is elevated(hyperglycemia), as after a meal, the pancreas receives signals to
release insulin into the blood.
Among its actions, insulin:
- Facilitates glucose
transport into the cells, especially those in muscle;
- Promotes glycogenolysis;
- Inhibits gluconeogenesis.
Insulin’s main function is to reduce the amount
of glucose circulating in the blood. But it is also involved in protein and fat metabolism, promoting cellular uptake of amino acids and
enhancing synthesis of protein and fat.
The pancreas secretes glucagon when the plasma glucose concentration falls below normal
concentrations(hypoglycemia). Its
effects generally oppose those of insulin. Glucagon promotes increased
breakdown of liver glycogen to
glucose(glycogenolysis) and increased gluconeogenesis,
both of which increase plasma glucose levels.
During exercise lasting 30 min or longer, the
body attempts to maintain plasma glucose concentrations. However, insulin
concentrations tend to decline. Research has shown that the ability of insulin
to bind to its receptors on muscle cells increases during exercise, due in large part to increased blood flow to
muscle. This increases the body’s sensitivity to insulin and reduces the need
to maintain high plasma insulin concentrations for transporting glucose into
the muscle cells. Plasma glucagons, on the other hand, shows a gradual increase
throughout exercise. Glucagon primarily maintains plasma glucose concentrations
by stimulating liver glycogenolysis. This increases glucose availability to the
cells, maintaining adequate plasma glucose concentrations to meet increased
metabolic demands. The responses of these hormones are usually blunted in
trained individuals, and those who are well trained are better able to maintain
plasma glucose concentrations.
“Physiology of sport and exercise”, fourth
edition; Jack H. Wilmore, David L. Costill, W. Larry Kenney
Hormones in human body - general info
Chemical classification of hormones
Hormones can be categorized as two basic types:
steroid hormones and nonsteroid hormones. Steroid
hormones have a chemical structure similar to cholesterol, since most
are derived from cholesterol. For this reason, they are soluble in lipids
and diffuse rather easily through cell membranes. This group includes the
hormones secreted by:
- The adrenal cortex(such as
cortisol and aldosterone);
- The ovaries(estrogen and
progesterone);
- The testes(testosterone),
and
- The placenta(estrogen and
progesterone).
Nonsteroid
hormones are not lipid
soluble, so they cannot easily cross cell membranes. The nonsteroid hormone
group can be subdivided into two groups: protein or peptide chormones and amino
acid-derived hormones. The two hormones from the thyroid gland(thyroxine and
triiodothyronine) and the two from the adrenal medulla(epinephrine and norepinephrine)
are amino acid hormones. All other nonsteroid hormones are protein or
peptide hormones.
Hormone actions
Because hormones travel in the blood, they
contact virtually all body tissues. How, then, do they limit their effects to
specific targets? This ability is attributable to the specific hormone
receptors possessed by the target tissues. The interaction between the hormone
and its specific receptor has been compared with a lock(receptor) and
key(hormone) arrangement, in which only the correct key can unlock a given
action within the cells. The combination of a hormone bound to its receptor is
referred to as a hormone-receptor complex.
Each cell typically has from 2,000 to 10,000
receptors. Receptors for nonsteroid hormones are located on the cell membrane, whereas those for
steroid hormones are found either in the cell’s cytoplasm or in its nucleus. Each
hormone is usually highly specific for a single type of receptor and binds only
with its specific receptors, thus affecting only tissues that contain those
specific receptors. Numerous mechanisms allow hormones to control the actions
of cells.
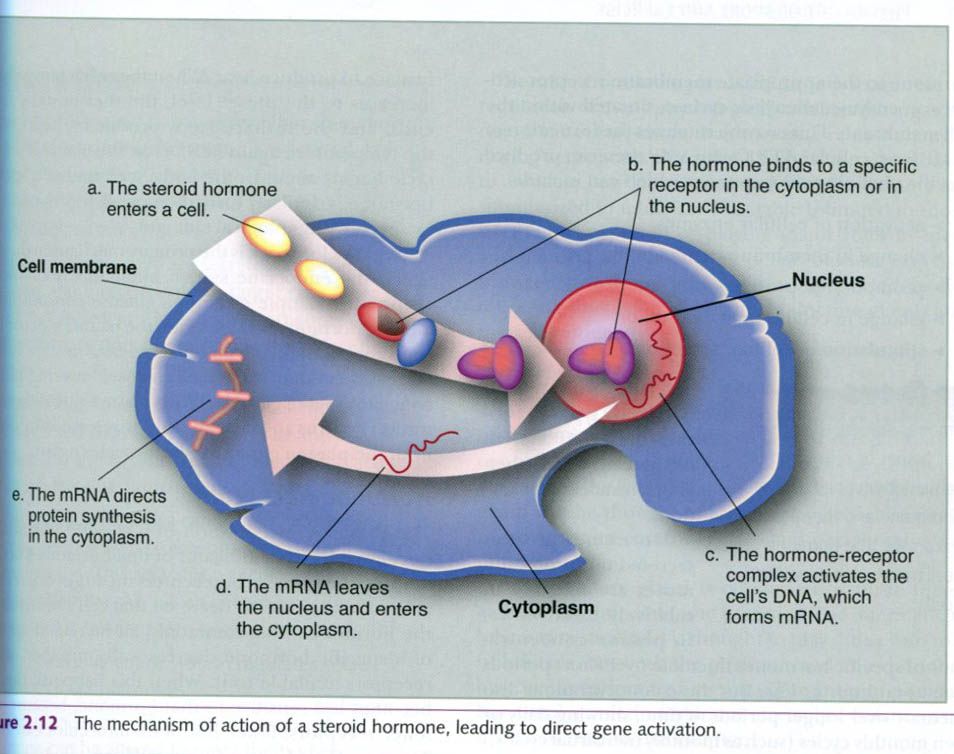
As mentioned earlier, steroid hormones are
lipid soluble and thus pass easily through the cell membrane. Their mechanism
of action is illustrated below. Once inside the cell, a steroid hormone binds
to its specific receptors. The hormone-receptor complex then enters the
nucleus, binds to part of the cell’s DNA, and activates certain genes. This
process is referred to as direct gene
activation. In response to this activation, mRNA is synthesized within the
nucleus. The mRNA then enters the cytoplasm and promotes protein synthesis. These
proteins may be:
- Enzymes that can have
numerous effects on cellular processes;
- Structural proteins to be
used for tissue growth and repair;
- Regulatory proteins that can
alter enzyme function.
Because nonsteroid hormones cannot cross the
cell membrane, they react with specific receptors outside the cell, on the cell
membrane. A nonsteroid hormone molecule binds to its receptor and triggers a
series of enzymatic reactions that lead to the formation of an intracellular second messenger. A widely distributed
second messenger that mediates a specific hormone-receptor response is cyclic adenosine monophosphate(cyclic AMP,
or cAMP). This mechanism is illustrated in the picture below.

In this case, attachment of the hormone to the
appropriate membrane receptor activates an enzyme, adenylate cyclase, situated
within the cell membrane. This enzyme catalyzes the formation of cAMP from
cellular ATP. Cyclic AMP then can produce specific physiological responses,
which can include:
- Activation of cellular
enzymes
- Change in membrane
permeability
- Change in cellular
metabolism
- Stimulation of cellular
secretions.
Thus, nonsteroid hormones typically activate
the cAMP system of the cell, which then alters intracellular functions.
Hormones are not secreted uniformly, but rather
are released in relatively brief bursts, so plasma concentrations of specific
hormones fluctuate over short periods such as an hour or less. But these
concentrations also fluctuate over longer periods of time, showing daily or
even monthly cycles(such as monthly menstrual cycles). How do endocrine glands
know when to release their hormones?
Most hormone secretion is regulated by a negative feedback system. Secretion
of a hormone causes some change in the body, and this change in turn inhibits
further hormone secretion. Consider how a home thermostat works. When the room
temperature decreases below some preset level, the thermostat signals the
furnace to produce heat. When the room temperature increases to the preset
level, the thermostat’s signal ends, and the furnace stops producing heat. When
the temperature again falls below the preset level, the cycle begins anew. In the
body, secretion of a specific hormone is similary turned on or off(or up or
down) by specific physiological changes.
Negative feedback is the primary mechanism
through which the endocrine system maintains homeostasis. Using the example of
plasma glucose concentrations and the hormone insulin, when the plasma glucose
concentration is high, the pancrease releases insulin. Insulin increases
cellular uptake of glucose, lowering plasma concentration of glucose. When
plasma glucose concentration returns to normal, insulin release is inhibited
until the plasma glucose level increases again.
Hormone receptors
The plasma concentration of a specific hormone
is not always the best indicator of that hormone’s activity because the number
of receptors on target cells can be altered to increase or decrease that cell’s
sensitivity to the hormone. Most commonly, an increased amount of a specific
hormone decreases the number of cell receptors available to it. When this
happens, the cell becomes less sensitive to that hormone, because with fewer
receptors, fewer hormone molecules can bind. This is reffered to as downregulation, or desensitization. In
some people with obesity, for example, the number of insulin receptors on their
cells appears to be reduced. Their bodies respond by increasing insulin
secretion from the pancreas, so their plasma insulin concentrations increase.
To obtain the same degree of plasma glucose control as normal, healthy people,
these individuals must release much more insulin.
In a few instances, a cell may respond to
the prolonged presence of large amounts of a hormone by increasing its number
of available receptors. When this happens, the cell becomes more sensitive to
that hormone because more can be bound at one time. This is reffered to as upregulation. In addition, one hormone
occasionally can regulate the receptors for another hormone.
Prostaglandins
Prostaglandins, although technically not hormones, are often
considered to be a third class of
hormones. These substances are derived from a fatty acid, arachidonic acid, and
they are associated with the plasma membranes of almost all body cells.
Prostaglandins typically act as local hormones, exerting their effects in the
immediate area where they are produced. But some also survive long enough to
circulate through the blood to affect distant tissues. Prostaglandin release
can be triggered by many stimuli, such as other hormones or a local injury.
Their functions are quite numerous because there are several different types of
prostaglandins. They often mediate the effects of other hormones. They are also
known to act directly on blood vessels, increasing vascular permeability(which
promotes swelling) and vasodilatation. In this capacity, they are important
mediators of the inflammatory response. They also sensitize the nerve endings
of pain fibers; thus, they promote both inflammation and pain.
“Physiology of sport and exercise”, fourth
edition; Jack H. Wilmore, David L. Costill, W. Larry Kenney
Пријавите се на:
Постови (Atom)











