



Respiratory adaptations to training
No matter how adept the cardiovascular system
is at supplying adequate amounts of blood to tissues, endurance would be
hindered if the respiratory system were not able to bring in enough oxygen to
meet oxygen demands. Respiratory system function does not usually limit
performance because ventilation can be increased to a much greater extent than
cardiovascular function. But, as with the cardiovascular system, the
respiratory system undergoes specific adaptations to endurance training to
maximize its efficiency.
After training, pulmonary ventilation is
essentially unchanged at rest. Although endurance training does not change the
structure of basic psychology of the lung, it does decrease ventilation during
submaximal exercise by as much as 20% to 30% at a given submaximal intensity.
Maximal pulmonary ventilation is substantially increased from a rate of about
100 to 120L/min in untrained sedentary individuals to about 130 to 150L/min or
more following endurance training. Pulmonary ventilation rates typically
increase to about 180L/min in highly trained athletes and can exceed 200L/min
in very large, highly trained endurance athletes. Two factors can account for
the increase in maximal pulmonary ventilation following training: increased
tidal volume and increased respiratory frequency at maximal exercise.
Ventilation is usually not considered a
limiting factor for endurance exercise performance. However, some evidence
suggests that at some point in a highly trained person’s adaptation, the
pulmonary system’s capacity for oxygen transport may not be able to meet the
demands of the limbs and the cardiovascular system. This results in what has
been termed exercise-induced arterial hypoxemia, in which arterial oxygen
saturation decreases below 96%. This desaturation in highly trained athletes
likely results from the large right heart cardiac output directed to the lung during exercise and consequently a decrease in
the time the blood spends in the lung.
Pulmonary diffusion, or gas exchange occurring
in the alveoli, is unaltered at rest and at standardized submaximal exercise
intensities following training. However, it increases during maximal exercise.
Pulmonary blood flow(blood coming from the heart to the lungs) appears to
increase following training, particularly the flow to the upper regions of the
lungs when a person is sitting or standing. This increases lung perfusion. More
blood is brought into the lungs for gas exchange, and at the same time
ventilation increases to that more air is brought into the lungs. This means
that more alveoli will be involved in pulmonary diffusion. The net result is
that pulmonary diffusion increases.
The oxygen content of arterial blood changes
very little with endurance training. Even though total hemoglobin is increased,
the amount of hemoglobin per unit of blood is the same or even slightly
reduced. The (a-ṽ)O2 difference, however, does increase with
training, particularly at maximal exercise intensity. This increase results
from a lower mixed venous oxygen content, which means that the blood returning
to the heart(which is a mixture of venous blood from all body parts, not just
the active tissues) contains less oxygen than it would in an untrained person.
This reflects both greater oxygen extraction at the tissue level and a more
effective distribution of blood flow to active tissue. The increased extraction
results in part from an increase in oxidative capacity of active muscle fibers.
In summary, the respiratory system is quite
adept at bringing adequate oxygen into the body. For this reason, the
respiratory system seldom limits endurance performance. Not surprisingly, the
major training adaptations noted in the respiratory system are apparent mainly
during maximal exercise, when all systems are being maximally stressed.
Adaptations in muscle
Repeated use of muscle fibers with endurance
training stimulates changes in their structure and function. Our main interest
here is in aerobic training and the changes it produces in muscle fiber type,
mitochondrial function, and oxidative enzymes.
Muscle
fiber types
Aerobic activities such as jogging and low-to
moderate-intensity cycling rely extensively on the slow-twitch(type I) fibers. In response to aerobic training, type I
fibers become larger. That is, they develop a larger cross-sectional area,
although the magnitude of change depends on the intensity and duration of each
training bout and the length of the training program. Increases of up to 25%
have been reported. Fast-twitch(type II) fibers, because they are not being recruited to the same extent, generally
do not increase cross-sectional area.
Most early studies showed no change in the
percentage of type I and type II fibers following aerobic training, but subtle
changes were noted among type II fiber subtypes. Type IIx fibers are used less
often than IIa fibers, and for that reason they have a lower aerobic capacity.
Long-duration exercise may eventually recruit these fibers into action,
demanding them to perform in a manner normally expected on the IIa fibers. This
can cause some IIx fibers to take on the characteristics of the more oxidative
IIa fibers. Recent evidence suggests that not only is there a transition of
type IIx to IIa fibers, but there can also be a transition of type II to type I
fibers. The magnitude of change is generally small, not more than a few
percentage points. As an example, in the HERITAGE Family Study, a 20-week
program of aerobic training increased type I fibers from 43.2% pretraining to
46.7% posttraining and decreased type IIx fibers from 20.0% to 15.1%, with type
IIa remaining essentially unchanged. These more recent studies have included
larger numbers of subjects and have taken advantage of improved measurement
technology; both might explain why changes are now being recognized.
Capillary
supply
One of the most important adaptations to
aerobic training is an increase in the number of capillaries surrounding each
muscle fiber. Endurance-trained men have considerably more capillaries in their
leg muscles than sedentary individuals. With long periods of aerobic training,
the number of capillaries has been shown to increase by more than 15%. Having
more capillaries allows greater exchange of gases, heat, wastes, and nutrients between the blood ad working muscle fibers. In fact, the increase in capillary density(i.e.
increase in capillaries per muscle fiber) is potentially one of the most
important alterations in response to training that allows the increase in VO2max.
It is now clear that the diffusion of oxygen from the capillary to the
mitochondria is a major factor limiting the maximal rate of oxygen consumption.
Increasing capillary density facilitates this diffusion, thus maintaining an
environment well suited to energy prediction and repeated muscle contractions.
Myoglobin
content
When oxygen enters the muscle fiber, it binds
to myoglobin, a compound similar to
hemoglobin. This iron-containing compound shuttles the oxygen molecules from
the cell membrane to the mitochondria. The
type I fibers contain large quantities of myoglobin, which gives these
fibers their red appearance(myoglobin is a pigment that turns red when bound to
oxygen). The type II fibers , on the
other hand, are highly glycolitic, so they require(and have) little
myoglobin-hence their whiter appearance. More important, their limited
myoglobin supply limits their oxygen capacity, resulting in poor aerobic
endurance for these fibers.
Myoglobin stores oxygen and releases it to the
mitochondria when oxygen becomes limited during muscle action. This oxygen
reserve is used during the transition form rest to exercise, providing oxygen
to the mitochondria during the lag between the beginning of exercise and the
increased cardiovascular delivery of oxygen.
Myoglobin’s precise contributions to oxygen
delivery are not yet fully understood. But aerobic training has been shown to
increase muscle myoglobin content to 75% to 80%. This adaptation would be
expected only if myoglobin enhances a muscle’s capacity for oxidative
metabolism.
Mitochondrial
function
Aerobic(oxidative) energy production takes
place in the mitochondria. Not surprisingly, then, aerobic training also
induces changes in mitochondrial function that improve the muscle fibers’
capacity to produce ATP. The ability to use oxygen and produce ATP via
oxidation depends on the number and size of the muscle mitochondria. Both
increase with aerobic training.
During one study that involved endurance
training in rats, the actual number of mitochondria increased approximately 15%
during 27 weeks of exercise. Average mitochondrial size also increased by about
35% over that training period. As the volume of aerobc training increases, so
do the number and size of the mitochondria.
Oxidative
enzymes
Regular endurance exercise has been shown to
induce major adaptations in skeletal muscle, including an increase in the
number and size of the muscle fiber mitochondrial capacity. The oxidative
breakdown of fuels and the ultimate production of ATP depend on the action of
mitochondrial oxidative enzymes, the special proteins that catalyze(i.e., speed
up) the breakdown of nutrients to form ATP. Aerobic training increases the
activity of these important enzymes.
Figure below illustrates the changes in the
activity of succinate dehydrogenase(SDH), a key muscle oxidative enzyme, over
seven months of gradually increased swim training. While the increases in VO2max
leveled off after the first two months of training, activity of this key
oxidative enzyme continued to increase throughout the entire training period.
This suggests that VO2max might be more influenced by the
circulatory’s system limitations with respect to transporting oxygen than by
the muscles’ oxidative potential.

The activities of muscle enzymes such as SDH
and citrate synthase are dramatically influenced by aerobic training. This is
seen in figure below, which compares the activities of these enzymes in untrained
people, moderately trained joggers, and highly trained runners. Even moderate
amounts of daily exercise increase these enzyme activities and thus the muscles
aerobic capacity. For example, jogging or cycling for as little as 20 min per
day has been shown to increase SDH activity in leg muscles by more than 25%.
Training more vigorously, for example for 60 to 90 min per day, produces a
two-to threefold increase in this activity.


One metabolic consequence of mitochondrial
changes induced by aerobic training is glycogen sparing, a slower rate of
utilization of muscle glycogen and enhanced reliance on fat as fuel source at a
given exercise intensity. This increase in the oxidative enzymes with aerobic
training most likely improves the ability to sustain a higher exercise
intensity, such as maintaining a faster race pace in a 10km run.
Metabolic adaptations to training
Three important variables are related to
metabolism:
- Lactate threshold
- Respiratory exchange ratio
- Oxygen consumption.
Lactate
threshold
Lactate threshold is a psychological marker
that is closely associated with aerobic endurance performance – the higher the
lactate threshold, the better the aerobic performance. Figure a below
illustrates the difference in lactate threshold that would occur following a 6
to 12-month program of endurance training. In either case, in the trained
state, one can exercise at a higher percentage of one’s VO2max before lactate
begins to accumulate in the blood. Because race pace in aerobic endurance
events is closely associated with lactate threshold, this translates into a
much faster race pace(see figure b). The reduction in lactate values at a given
rate of work to likely attributable to a combination of reduced lactate
production and increased lactate clearance.

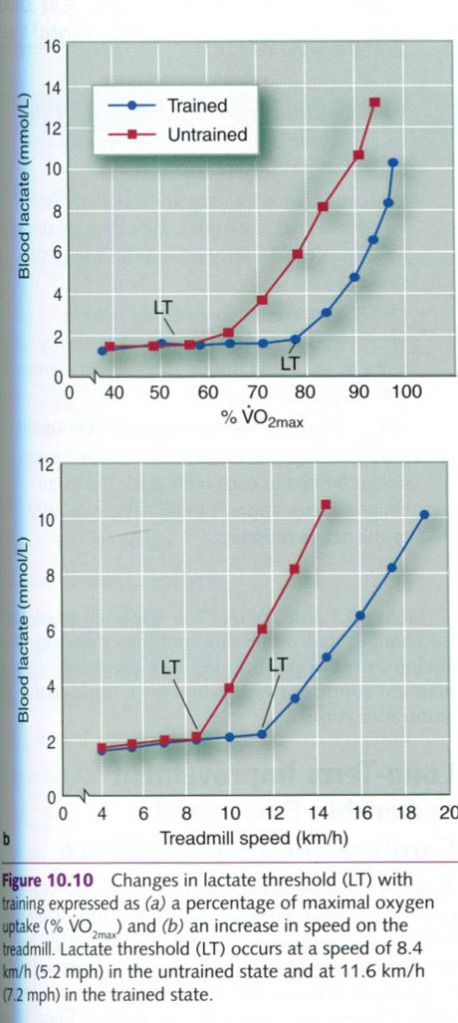
The concentration of lactate in the blood
following a fixed-pace swim or run provides an excellent means of monitoring
the physiological changes that occur with training. As athletes become better
trained, their blood lactate concentrations are lower for the same rate of
work. This suggests that they are developing greater aerobic power, a reduced
reliance on the glycolytic system for energy, or perhaps both.
The respiratory
exchange ratio(RER) is the ratio of carbon dioxide released to oxygen
consumed during metabolism. The RER reflects the type of substrates being used
as an energy source.
After training, the RER decreases at both
absolute and relative submaximal exercise intensities. These changes are
attributable to a greater utilization of free fatty acids instead of carbohydrate at these work rates
following training.
Resting
and submaximal oxygen consumption
Oxygen consumption(VO2) at rest is
unchanged following endurance training. While a few cross-sectional comparisons
have suggested that training elevates resting VO2, the HERITAGE
FAMILY STUDY – with a large number of subjects and with duplicate measures of
resting metabolic rate both before and after 20 weeks of training – showed no
evidence of an increased resting metabolic rate after training.
During submaximal exercise at a given exercise
intensity, VO2 is either unchanged or slightly reduced following
training. In the HERITAGE Family Study, with more than 700 participants,
training reduced submaximal VO2 by 3.5% at a work rate of 50W. There
was a corresponding reduction in cardiac output at 50W, reinforcing the strong
interrelationship between VO2 and cardiac output. A decrease in VO2
during submaximal exercise could result from an increase in exercise economy(performing
the same exercise intensity with less extraneous movement).
Maximal
oxygen consumption
VO2max is the best indicator of
cardiorespiratory endurance capacity and increases substantially in response to
endurance training. While small and very large increases have been reported, an
increase of 15% to 20% is typical for a previously sedentary person who trains
at 50% to 85% of his or her VO2max three to five times per week, 20
to 60 min per day, for six months. For example, the VO2max of a
sedentary individual could reasonably increase from a 35ml x kg-1 x
min-1 to 42ml x kg-1 x min-1 as a result of
such a program. This is far below the values we see in world-class endurance
athletes, whose values generally range from 70 to 94ml x kg-1 x min-1.
The more untrained an individual is when starting an exercise program, the
larger the increase in VO2max.












1 коментара:
Thật quá tuyệt vời vì những nội dung trong bài của bạn, tôi cho rằng là sẽ có rất nhiều người tìm những nội dung này
mình cũng xin góp thêm 1 chút ý kiến ở trong link sau đây: áo quần tập aerobic
Постави коментар